热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
背景介绍:完全淹没的植物依靠与周围水的气体交换,而被淹没的沉积物通常在表层以下几毫米处就缺氧,但其中的CO2含量却很高。在白天沉水植物通过充满气体的通气组织将幼苗产生的氧气进行光合作用,从而维持地下组织的有氧呼吸。低潮有时会导致水量减少,如果在深海草丛中形成密集冠层的系统中夜间发生低潮,则除了沉积物中O2消耗外,大量呼吸性生物质可能会消耗含有O2的水并导致海草根茎和根中的严重的缺氧相比之下,当白天出现低潮时,由于大量的光合作用生物量,溶解的O2可以增加,但是如果发生热胁迫,太阳辐射可能会很快加热浅水,导致净光合作用(PN)降低。也增加了光呼吸,因为当二氧化碳耗尽而产生氧气时,O2:CO2比率会增加。海草通常永久性地浸没在海水中,而大量溶解有O2的水通常可以防止水柱中的严重缺氧。在本论文研究中,研究人员测量了两种热带海草物种泰来藻和海菖蒲在田间位置占主导地位的内部组织通气,以及以潮汐和海浪为特征的热带浅潮海草草甸的关键环境变量。真实的昼夜温度差和O2振幅。假设在夜间低潮会导致水体缺氧和组织缺氧,而在白天低潮时,即使热胁迫导致叶片PN下降(而非光合组织RD增加)。在实验室实验中,测试了温度如何影响两个物种的叶片的PN和RD,从而使我们能够比较它们的生理学。
实验室中应用unisesne微呼吸系统测量了叶片段的水下净光合作用(PN)和暗呼吸作用(RD)。首先将组织样品在封闭的透明小瓶中,在温和混合的条件下,在确定的培养基中孵育已知的时间,并在恒温(适当的情况下,在光源或黑暗中下保持恒温。O2的释放量(P N)或消耗量(RD)是通过使用unisense氧气微电极测量瓶子内叶片组织获得的,从两个物种的每一个中取了五片重复的叶子。从叶片中间三分之一处切出5(PN)至10(RD)mm长的叶段(投影面积约30-150 mm2)。根据Pedersen描述的光合有效辐射方法,在25-50℃、间隔5℃的条件下,使用25 ml的玻璃小瓶,以确保按照Pedersen等人的方法旋转小瓶时进行混合。在两个小瓶内使用强度为760±76μmol光子m-2 s-1(平均值±SE,n=4),该设备是具有光合作用活性辐射装置(PAR),这是由两个水平放置的金属卤化物灯。孵化介质为0.7µm过滤海水,盐度为35‰,初始pO2接近空气平衡,没有叶段的小瓶用作空白。在已知孵育时间(90-150分钟)后,使用unisense O2微型氧电极(OX-500,Unisense A/S,丹麦)连接到一个1通道皮安培表。通过O2动力学原位测量的O2的动态消耗,从而算出叶片在光辐射下对应的O2的释放量(P N)或消耗量(RD)。
实验结果
极端的大潮环境对礁滩上同时存在的两种海草物种提出了挑战。白天浅水的特点是高PO2,pH和温度影响PN和R d。Pn下降是由于热应力和CO 2可用性接近零,而R d在温水增加。在夜间,通过对比,溶氧下降,从而导致在两个拍摄基地T.hemprichii和E.acoroides缺氧小时低氧水平。研究证明了PN从前一天暴露于40°C到前一天的4小时,这两个物种的R和R D都在夜间完全恢复,而45°C的热应激导致组织永久受损。这两种海草生长在曝气海水中,但通常生长在低氧沉积物中,升高温度和低氧是应激因素。这些海草“生活在边缘地带”,能够忍受当前的diel O2和极端温度,但如果温度上升,这两种海草在这个栖息地都可能受到威胁。
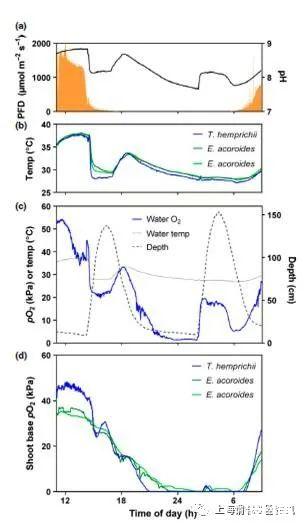
图1、具有关键环境参数的一昼夜循环示例:沉积物上方10 cm处的水下光子通量密度(PFD)和环境水柱中的pH(a),三株复制植物的芽基旁边的温度(b),水柱pO2,温度和深度10厘米以上的沉积物(c)和原位O2代动力学中的拍摄基地。泰来藻(1种植物)和海菖蒲属(2种植物)。(d)在部署后不久,由于光电极被移动,血淋巴细胞的一个复制体丢失了。之所以选择这个例子,是因为带着含海水的O2进入潮汐的影响与日出后光合作用产生的O2明显分离。
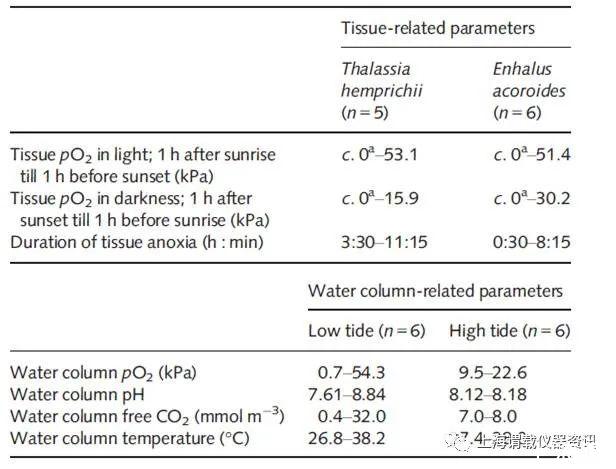
图2、除主要环境参数的波动(水柱pO2)之外,两种热带海草物种泰来藻和海菖蒲属在明亮和黑暗以及严重组织缺氧或缺氧的持续时间下芽基的原位组织pO2,pH和温度)在三个昼夜时期的现场情况下在西澳大利亚塔隆岛的热带礁石上实测。
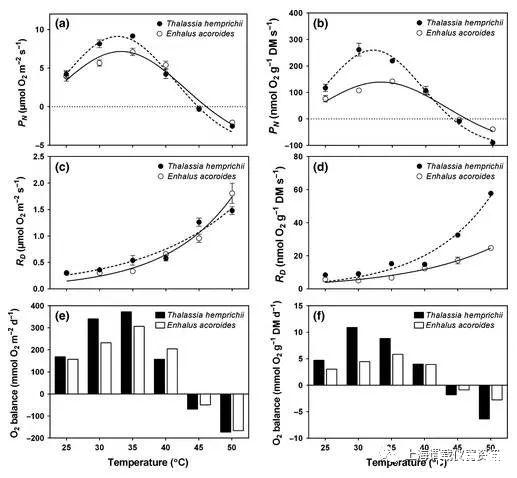
图3、两种热带海草物种Thalasia hemprichii和Enhalus acoroides的净水下光合作用(P N;a,b),暗呼吸(R D;c,d)和估计昼夜间伐叶水平氧气平衡(e,f)与温度的关系。速率表示为单位投影表面积(a,c,e)和单位质量(b,d,f)。高斯模型用于估计(a)和(b)中的最佳温度(T opt)和最大光合作用(P max)对于每个温度,O 2假设在12小时的光照和12小时的暗处与野外情况相似,则计算出平衡(图1)。(a–d)中的值为平均值±SE(n=4-5)。
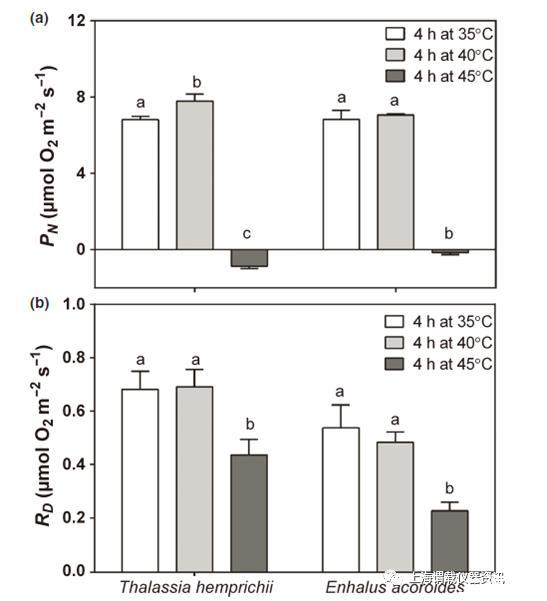
图4、两种热带海草物种,Thalassia hemprichii和海菖蒲属水下净光合(PN,a)和暗呼吸(R d,b)。将整株植物在35、40或45°C的光照下暴露4 h,然后在25°C的黑暗中恢复过夜,然后在12–18 h进行35°C的测量。不同的字母表示每种物种之间的显着差异。

图5、碳酸氢盐的使用抑制作为无机碳的水下净光合源(PN)的两种热带海草物种,Thalassia hemprichii和海菖蒲属,使用TRIS(50molm-3)添加至海水和pH调节至8.00,以防止酸化细胞壁空间和扩散边界层的分布,与海水中的对照相比。使用了海水的“pH漂移实验”作为碳酸氢盐使用的诊断测试,这表明这两种物种均能够在35°C的温度下利用碳酸氢盐作为无机碳源。
结论与展望
本论文主要对两个热带海草(Thalasia hemprichii和Enhalus acoroides)中测量内部通气进行了相关研究,研究发现它们会随着极端潮汐和昼夜的温度幅度的增长而增长。通过使用unisense微电极系统研究了温度对叶片净光合速率(PN)和暗呼吸速率(RD)的影响。浅水池白天低潮时pO2高(54 kPa),pH(8.8),温度(38℃)。这是由于PN在33°C时最大,所以高温和CO2降低会降低PN,而RD(Q 102.0-2.7的Q 10)高于33°C时(分别为0.45和0.33μmolO2 m-2 s-1)会增加。在夜间退潮时,O2下降,导致两个物种的芽基缺氧,但流入的水中含有碳。暴露于40°C环境下4 h的海草的芽显示P N和R D的恢复,而45°C则导致了叶片受损。这些研究结果突显了这两个物种对高温的相似耐受性以及浅水池中海草的热胁迫威胁,也说明了这些礁滩上的海草“生活在其生理耐受性的边缘”。
 相关新闻
相关新闻