热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
微生物腐蚀是金属劣化的一种常见形式,会对许多工业应用产生负面影响,包括石油管道、海上平台和船舶、核电站设施、医疗仪器,甚至空间站。目前含铁金属的微生物腐蚀模型表明,微生物主要通过间接机制在需氧条件下增强腐蚀,而厌氧腐蚀通常与厌氧呼吸更直接相关。希瓦氏菌MR-1是一种有吸引力的模型微生物,用于阐明每年对工业应用造成数十亿美元腐蚀损害的生物膜-金属相互作用。已经提出了沙雷菌增强腐蚀的多种机制,但这些机制以前都没有用排除电子转移替代途径的方法进行严格研究。研究发现在有氧条件下生长的希瓦氏菌在不锈钢试样上形成厚的生物膜(~50µm),加速了对无菌对照的腐蚀。H2和核黄素被排除在中间电子载体之外,因为不锈钢没有减少核黄素,并且先前的研究表明不锈钢不会产生H2。菌株ΔmtrCBA,其中希瓦氏菌中最丰富的孔蛋白-细胞色素导管的基因被删除,在需氧培养中腐蚀不锈钢的程度大大低于野生型。野生型生物膜在厌氧条件下以不锈钢作为唯一电子供体很容易还原硝酸盐,但菌株ΔmtrCBA没有。这些结果表明希瓦氏菌可以直接消耗含铁金属的电子,并说明金属到微生物的直接电子转移如何成为腐蚀的重要途径,即使在有氧环境中也是如此。
Unisense 氧气微电极系统的应用
沙雷菌生物膜在LB琼脂板上在30°C下生长两天,用于测量生物膜下方的氧气浓度。使用尖端直径为5μm的Unisense氧气微传感器(OX-5,Unisense A/S,Aarhus,Denmark)。传感器的测量步径为3μm,测量周期为1秒,测量之间的等待时间为3秒。SensorTrace Profiling软件用于分析数据。
实验结果
本论文研究了沙雷菌腐蚀不锈钢的机制,不锈钢是一种用于制造各种结构和装置的含铁金属。提供了通过孔蛋白-细胞色素复合物直接吸收电子的证据,并证明在好氧环境中生长的微生物可以通过直接的金属与微生物的电子交换来腐蚀金属,这一发现对设计缓蚀策略具有重要意义。
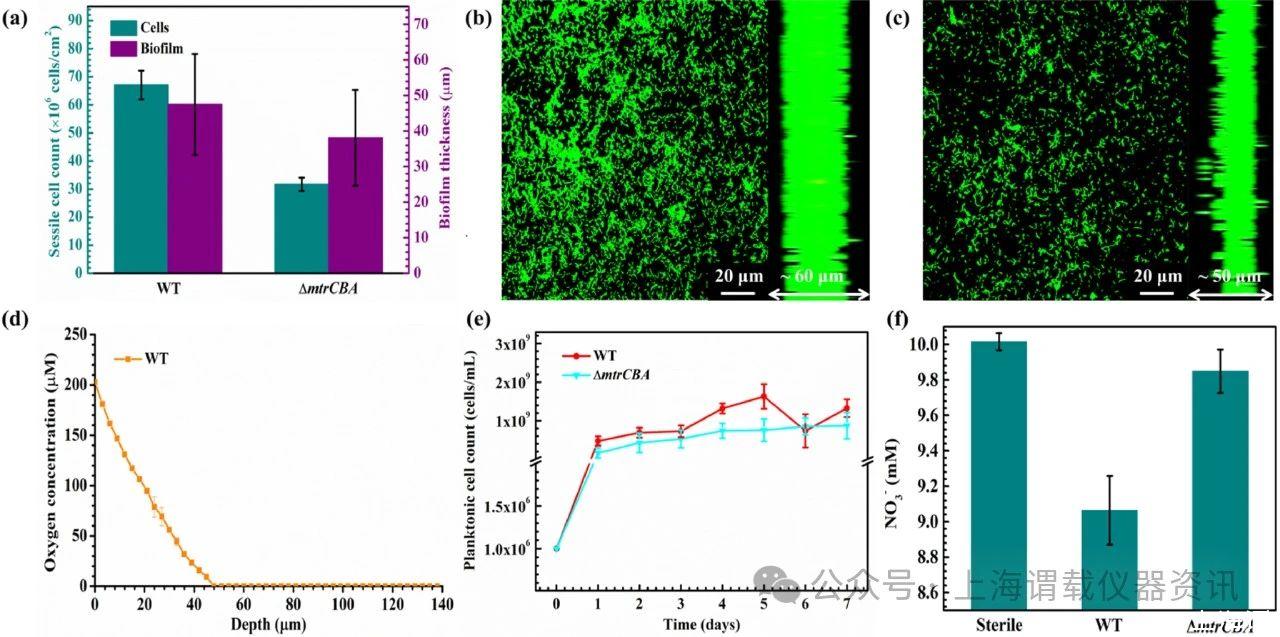
图1、沙雷菌菌株的生物膜、固着细胞计数、氧浓度、浮游细胞计数和NO 3-浓度。a孵育7天后316 L SS试样表面上的固着细胞计数和生物膜厚度。b野生型S.oneidensis MR-1菌株的CLSM图像。cΔmtrCBA应变的CLSM图像。d野生型沙雷菌MR-1菌株生物膜层中的氧气浓度。e使用316 L SS试样在整个7天孵育期间的浮游细胞计数。f NO 3-使用316 L SS试样在整个7天孵育过程中的浓度。
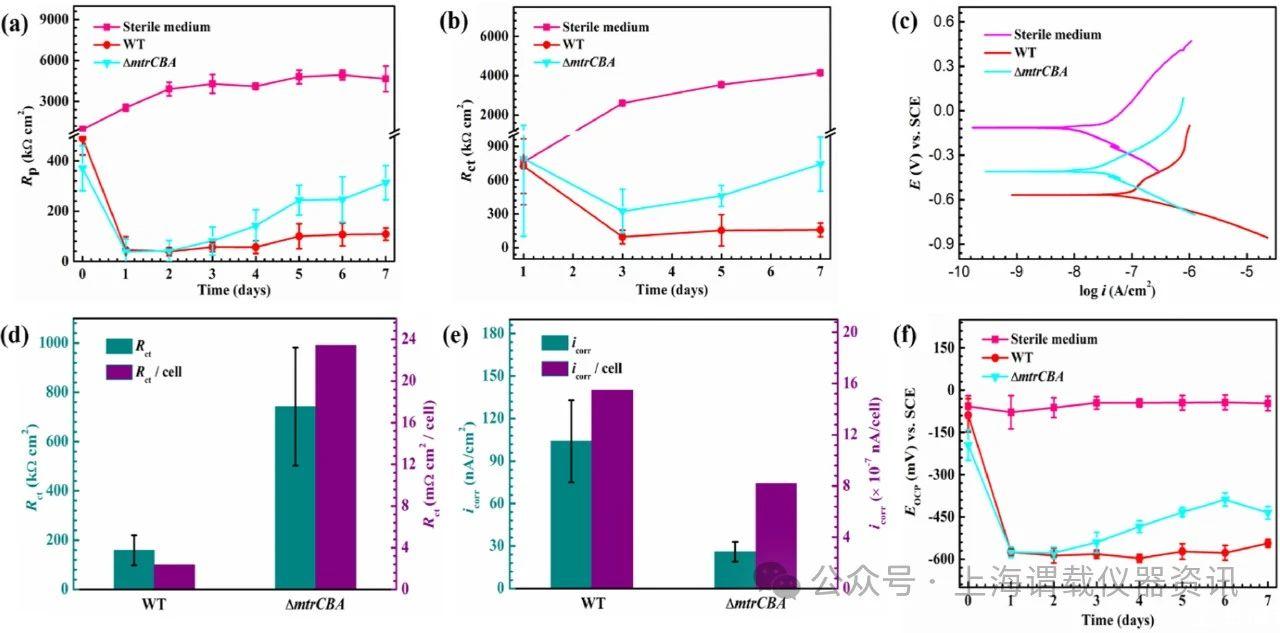
图2、各种沙雷菌菌株的腐蚀速率和归一化腐蚀速率。在无菌LB培养基或含有两种沙雷菌菌株之一的培养基中培养的316 L SS试样随时间的R p数据。b)R ct随时间的变化。c)培养7天后测量的动电位极化曲线。d从EIS测量中获得的R ct数据以及在孵育7天后针对工作电极上的固着细胞数标准化的R ct。e)腐蚀电流密度(i corr)从动电位极化曲线中获得,i)对固着细胞数进行了归一化。f)在有或没有各种沙雷菌菌株的LB培养基中孵育的316 L SS试样的E OCP随时间的变化。误差线代表不同电化学玻璃电池中至少三个独立试样的标准偏差。
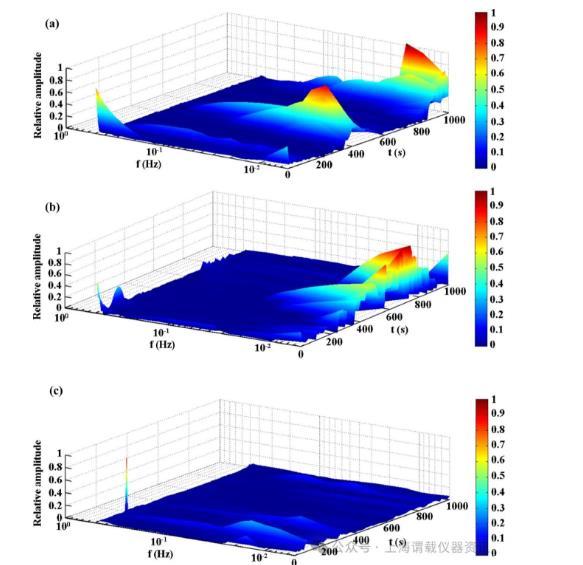
图3、浸泡7天后的代表性希尔伯特光谱。a在无菌LB培养基中。b在野生型S.oneidensis MR-1菌株接种培养基中第7天。c在ΔmtrCBA菌株接种培养基中。
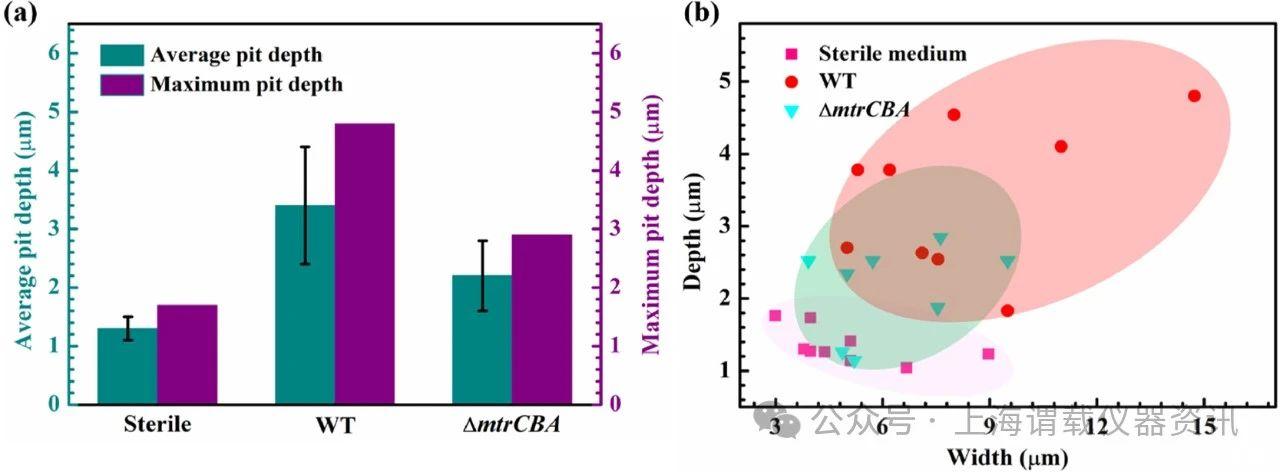
图4、由沙雷菌生物膜引起的点蚀。a浸入无菌LB培养基或接种两种沙雷菌菌株之一的LB培养基7天后,316 L SS试样表面的最大和平均凹坑深度。b浸入无菌LB培养基或接种两种沙雷菌菌株之一的LB培养基后,在316 L SS试样表面上观察到的凹坑的深度和宽度。
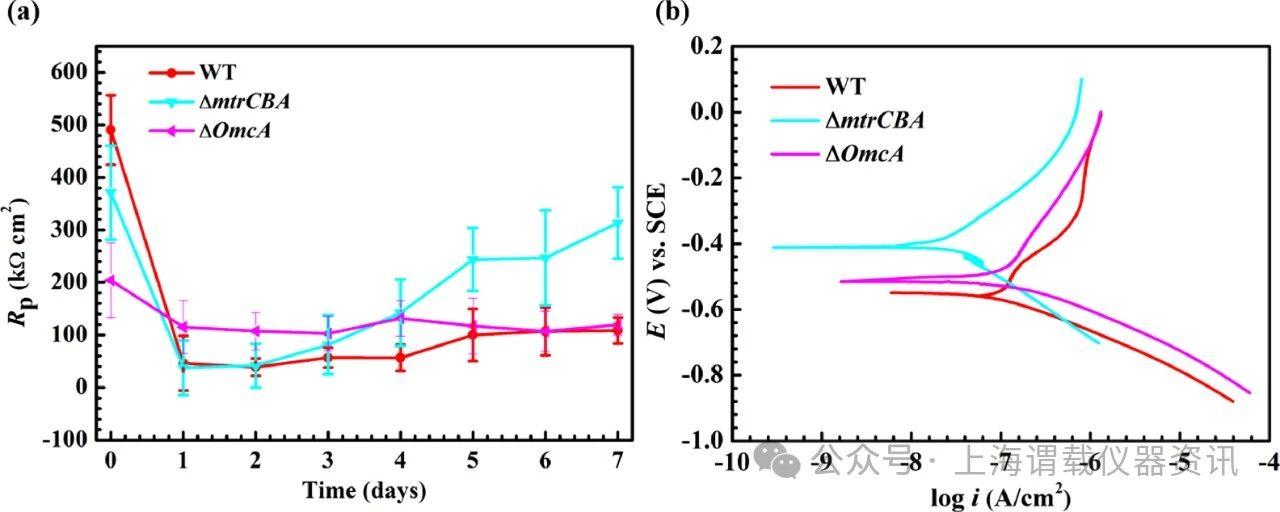
图5、用316 L SS试样孵育7天后,对沙雷菌的OmcA缺陷菌株进行电化学分析。a在具有316 L SS试样的LB培养基中孵育不同的沙雷菌菌株后,R p随时间的变化,b孵育7天后测量的动电位极化曲线。
结论与展望
在这项研究中,研究人员使用跨学科方法来证明通过孔蛋白细胞色素导管直接吸收电子,这是一种通常与无氧呼吸相关的细胞外电子交换机制,可能是有氧环境中不锈钢腐蚀的重要机制。直接电子吸收机制与先前记录的有氧条件下的腐蚀途径明显不同。研究发现了一种以前未被认识的有氧条件下金属腐蚀的途径,为金属腐蚀生物膜的生态学提供了新的见解,这可能对指导新的腐蚀缓解策略的发展很重要。
 相关新闻
相关新闻