热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
导读
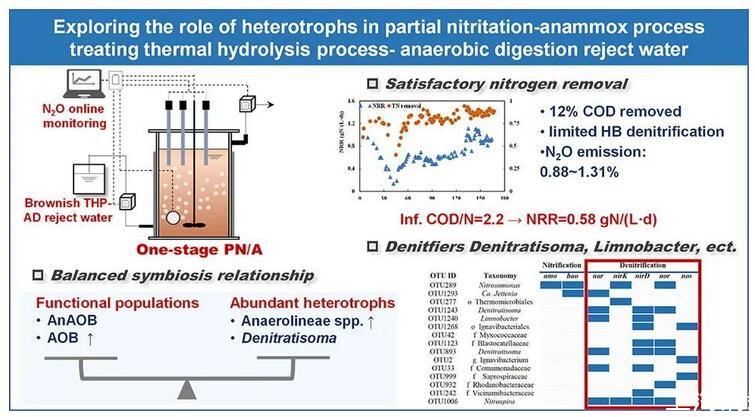
异养细菌(HB)普遍存在于厌氧氨氧化工艺中,但其在部分硝化-厌氧氨氧化(PN/A)工艺处理高有机物废水中的功能和生态作用仍未清楚。本研究旨在阐明单级PN/A处理热水解工艺(THP)-厌氧消化(AD)废水中的HB活性及其微生物相互作用。PN/A反应器实现了令人满意的氮去除率,高达0.58±0.06 g N/(L·d),同时THP-AD废水中的COD去除率约12%。PN/A反应器的N2O排放因子为1.15%±0.18%(处理合成废水)和0.95%±0.06%(处理消化液)。在反应器运行过程中,HB和功能微生物(即厌氧氨氧化细菌和好氧氨氧化细菌)之间保持着平衡的共生关系。Anaerolineae spp.的相对丰度明显增加,而反硝化菌Denitratisoma在处理THP-AD废水时略有减少。异养生物对电子供体的偏好解释了增长趋势的差异。
实验设计与方法
结果与讨论
1 PN/A工艺性能1.1反应器性能
PN/A反应器已在稳定脱氮性能下运行170多天(表1)。由于接种厌氧氨氧化颗粒污泥中的AOB丰度低,因此在启动阶段I时优化曝气和进水NH4+方案以刺激AOB的生长。间歇曝气是PN/A运行期间有效的曝气策略,本研究中通过定期打开和关闭空气供应来采用(表1)。进水NH4+浓度在63天内从124增加到243 mgN/L,而亚硝酸盐仅在前30天供应。运行60天后,出水NH4+浓度低于2 mgN/L(图1,A),实现了有效的NH4+去除,表明PN/A工艺完成。NO3-生成与总脱氮比(ΔNO3-/ΔTN)反映了该工艺的生物氮转化,仅考虑AOB和AnAOB活性的情况下,理论ΔNO3-/ΔTN为0.13。在第一阶段,ΔNO3-/ΔTN比值在0.15~0.48的范围内变化,高于理论值,表明可能有亚硝酸盐氧化细菌(NOB)产生NO3-。随着NLR的降低,NRR从前32天(有亚硝酸盐供应)的0.94±0.39 gN/(L·d)下降到第52~63天(无亚硝酸盐供应)的0.53±0.08 gN/(L·d)(图1,B)。
在第二阶段(处理RW),进水从SW变为THP-AD废水。进水COD浓度为547±16 mgCOD/L,COD/NH4+-N比率为2.2±0.1。出水COD浓度为484±52 mgCOD/L,COD去除率约为12%。COD去除率相对低于以往研究(两阶段PN/A系统实现了50.4%到80%的高COD去除率,而PN反应器去除了53%到66%的COD。可能的解释可能为:(1)与之前的两阶段PN/A系统的PN反应器(0.8~2 mgDO/L)相比,本研究中的DO(<0.05 mg/L)较低,好氧阶段的氧气限制导致COD氧化效果较差。(2)与之前的研究相比,THP-AD废水中有机物的生物降解性较低。有机物的可生物降解性高度决定了于厌氧消化效率和稳定性,因此不同污水源的有机物特性不同是合理的。从第74天到第101天,出水NO2-浓度保持相对较低。同时,出水NO3-显著减少,导致ΔNO3-/ΔTN比I期低0.16。RW处理期间,体系实现了令人满意的NRR(0.58±0.06 gN/(L·d)),与之前实验室规模和中试规模的研究相当。
由于从第103天开始出现NO2-积累,在第三阶段(后RW处理)将进水改为合成废水。此后,NO2-积累从第132天开始消失。ΔNO3-/ΔTN比值下降到0.09,NRR大大增加到0.91 gN/(L·d)。亚硝酸盐积累表明NO2-生成率(主要由AOB完成)高于NO2-消耗率(主要由AnAOB完成)。尽管有报道称废水中不溶性有机物能对AOB产生抑制,但当注入THD-AD废水时,AOB仍然保持活性。如前所述,在进水切换回合成废水后,亚硝酸盐积累消失,这表明THP-AD废水可能抑制了AnAOB活性。
表1操作条件和反应器性能总结。
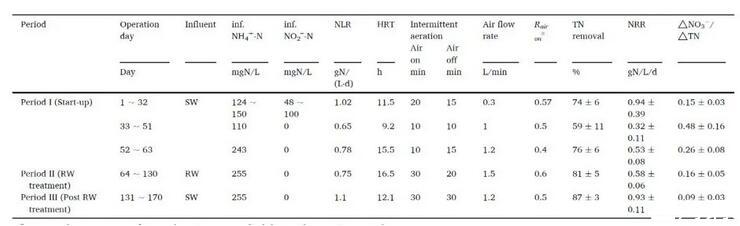
a R air on:反应期间供应空气的时间百分比。
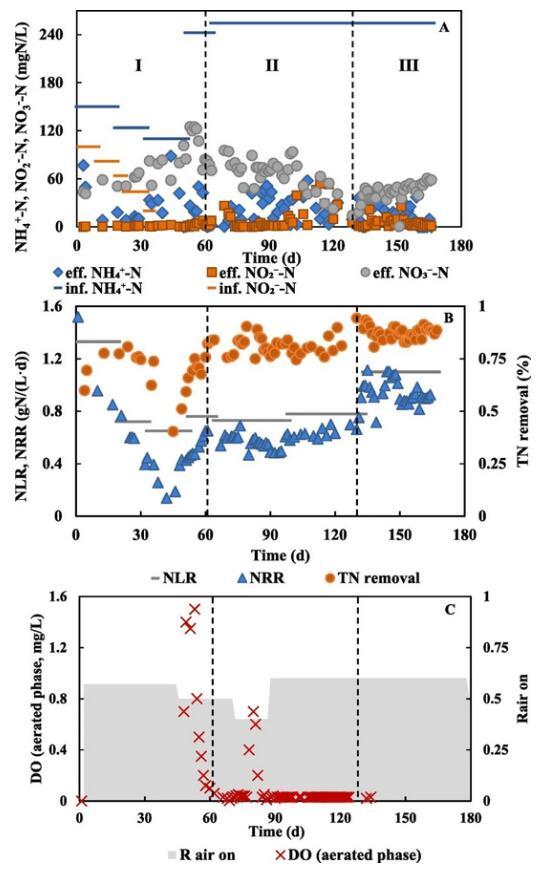
图1 PN/A反应器性能。A:进水和出水中的含氮物质。B:氮负荷和氮去除。C:Rair on(反应期间供应空气的时间百分比)和DO。
1.2细菌活性
为确定PN/A污泥在不同运行阶段的细菌氮转化活性,在反应器运行期间进行了批量实验测试厌氧氨氧化活性。在第一阶段取样的污泥显示出高厌氧氨氧化活性(SAA=851±8 mgN/(gVSS·d))。在第二阶段,合成废水中的SAA降至504±6 mgN/(gVSS·d),THP-AD废水中的SAA降至501±55 mgN/(gVSS·d)(图2)。在第三阶段,将进水改回合成废水后,SAA恢复到558.3 mgN/(gVSS·d)。厌氧氨氧化活性的演变表明,THP-AD废水可能会阻碍厌氧氨氧化活性。同样,之前的一项研究表明,THP-AD中的可溶性有机物(尤其是不可生物降解部分),直接抑制了AnAOB活性。
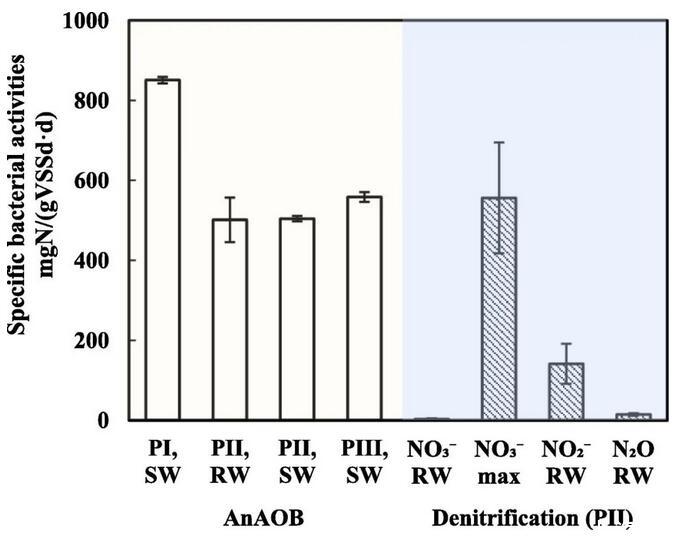
图2 AnAOB的特定细菌活性和异养反硝化作用。PI:第一期;PII:第二期;PIII:第三期;SW:合成废水;RW:消化液。
2 PN/A工艺的N2O排放概况
在合成废水处理(I期)和RW处理(II期)期间监测氮形态和N2O分布(图3)。虽然曝气阶段的空气流速为2 L/min,但观察到的DO较低(0.03 mgO2/L);在非曝气阶段,DO降至0.01 mgO2/L。循环开始时的NO2-浓度为2.8 mgN/L,最大NO2-为13.6 mgN/L。N2O浓度响应于曝气条件(图3,C)。在曝气阶段,N2O浓度因剥离而逐渐降低,而在非曝气阶段,N2O浓度逐渐增加。在I期,PN/A反应器的比净N2O产率为1.9±0.2 mgN2O-N/(gVSS·d),N2O排放因子(ΔN2O/ΔNH4+)为1.15%±0.18%。第二阶段氮形态显示出与第一阶段相似的模式,COD在整个循环中从544 mg/L下降到456 mg/L(图3,B)。DO大部分时间都保持在0.05 mg/L以下,而在最后一个曝气阶段,由于NH4+耗尽,DO增加到超过1 mg/L的水平。当反应器加入THP-AD废水时,N2O排放因子为0.95%±0.06%,略低于合成废水。
本研究的N2O排放因子(0.88~1.31%)与之前对不添加COD的一阶段PN/A工艺处理合成废水的研究和处理废水(1.2%~2%的进水TN)相当。PN/A工艺中N2O产生的来源主要包括:(1)异养反硝化菌的异养反硝化作用;(2)AOB硝化作用;(3)AOB的硝化反硝化作用。唯一的生物N2O消耗途径是异养反硝化菌的异养N2O还原。对于不添加COD的PN/A工艺,AOB被认为是N2O的主要生产者;而与AOB相比,HB在N2O产生中的作用似乎微不足道。相比之下,以往的研究表明,在存在可生物降解有机物时,异养N2O生产为普遍途径。因此,AOB可能是用合成废水供给时PN/A反应器中N2O的主要生产者,异养N2O途径可能在处理RW时得到促进。我们的结果表明,RW中的N2O排放量略低于合成水,因为异养N2O途径受到THP-AD废水的影响。Zhu和Chen(2011)报道了污泥RW降低了好氧-缺氧反应器的N2O和NO排放,推测污泥废水中的Cu2+和丙酸增加了N2O还原酶活性与NO还原酶活性的比率,导致N2O排放量减少。同时,废水中的有机物可能会改变异养N2O生成和消耗途径的平衡。据报道,废水的低COD/N(0.4)增加了硝化反应器中N2O的生成,而充足的有机物供应减少了好氧-缺氧反应器和PN/A反应器中N2O的排放。因此,异养N2O途径受THP-AD废水组分的影响。
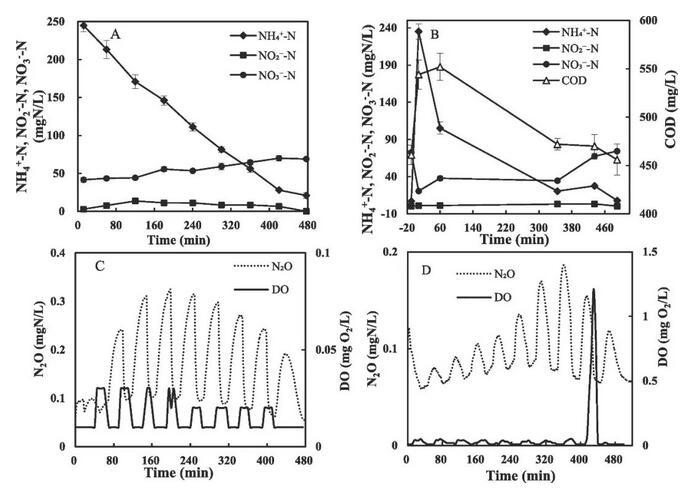
图3 SBR循环过程中PN/A反应器的氮形式和N2O排放分布。A、C:处理合成废水。B、D:处理THP-AD废水。
3 PN/A反应器微生物群落演替
从反应器运行周期和粒径等角度分析了微生物群落组成。16S rRNA测序平均每个样本产生了49,383个读取,序列聚类后产生1381个OTU,相似度为97%。通过在至少3个样本中设置高于5的OTU序列来过滤低丰度OTU。过滤后,剩余486个OTU用于进一步分析。在门水平上,绿藻门、变形菌门、拟杆菌门、浮游菌门和酸杆菌门为优势门。在属水平上,优势属包括c_Anaerolineae_1、f_Saprospiraceae_1、不动杆菌属、o_Kapabacteriales、Nitrosomonas、Denitratisoma和Candidatus Jettenia(混合污泥,图4,A)。Nitrosomonas spp.(唯一确定的AOB物种)的相对丰度随着反应器的运行而增加,其在接种物中的相对丰度相当低(0.7%),但在第166天达到3.2%(混合污泥,图4,A)。正如其好氧代谢所预期的那样,Nitrosomonas spp.在絮状污泥(直径<200μm)的丰度高于颗粒污泥(直径>200μm)(图4,B)。另一方面,Nitrospira spp.(NOB)的相对丰度保持在1%以下。因此,通过应用间歇曝气的方法,AOB在PN/A工艺中逐渐富集而没有NOB富集。我们的结果与之前的研究一致,表明间歇曝气是促进AOB生长和抑制NOB的有效方法。
Ca.Brocadia是接种物中的主要AnAOB物种(相对丰度7.8%),但在启动阶段和RW处理期后其相对丰度大大降低(低至0.1%)(混合污泥,图4,A)。在后RW处理期间,Ca.Brocadia的相对丰度在第166天逐渐增加,达到8.8%。相比之下,另一种AnAOB Ca.Jettenia也存在于接种物中(相对丰度2.1%),其丰度在RW处理期间增加,然后在RW后处理期间保持相对稳定(混合污泥,相对丰度4.1%)。观察到的Ca.Brocadia和Ca.Jettenia的差异增长趋势与AnAOB的生态位分化理论一致,Ca.Brocadia spp.是通常在高底物环境中发现的r策略者,而Ca.Jettenia spp.是更好地适应底物限制环境的k策略者。接种物富含厌氧氨氧化的污泥适应进水中足够的NH4+和NO2-,NLR高达3.5 gN/(L·d),适合Ca.Brocadia生长。相反地,PN/A工艺受NO2-限制,因为NO2-的产生取决于好氧NH4+氧化速率。这种低底物条件有利于k-策略者Ca.Jettenia的生长。此外,与絮状污泥中的Ca.Brocadia和Ca.Jettenia相比,颗粒污泥中二者丰度更高(图4,B)。
HB在PN/A污泥中高度多样化且含量丰富。Chloroflexi spp.在厌氧氨氧化工艺中广泛被观察到,其相对丰度从接种中的32±0%增加到RW处理期间的47±4%,然后在RW处理后进一步增加到54±8%。同时,c_Anaerolineae_1(隶属于Chloroflexi门)从接种时期到RW处理期(混合污泥,图4,A)的丰度由14±3%增加到24±6%,再进一步增加到33±8%。有趣的是,c_Anaerolineae_1在颗粒污泥中非常丰富(高达总群落的24%),而它们在絮状物中的存在是有限的(低于总群落的7%)。变形菌属也是普遍的种群,其相对丰度从接种物中的22±4%分别下降到RW处理期和后RW处理期的16±1%和15±2%。Denitratisoma隶属于Proteobacteria,其丰度从接种物中的6±1%分别略微下降到RW处理和后RW处理后的4±1%和4±1%(混合污泥,图4,A)。Limnobacter属于Proteobacteria,其在反应器运行过程中也逐渐减少。Denitratisoma和Limnobacter在颗粒污泥中均显示出更高的丰度(图4,B)。
以THP-AD废水作为进水,功能种群(AnAOB和AOB)不断增长并保持活跃,而HB种群则显示出不同的增长趋势。虽然HB和功能性群体对底物存在竞争,但HB和AnAOB之间的代谢相互依赖,例如氨基酸营养共生、亚硝酸盐循环或代谢物交换。功能菌群和HB可以在低浓度有机废水中共存。先前在两阶段PN/A系统中处理COD/NH4+-N为0.97的废水的研究表明,Ca.Kuenenia属保持相对稳定,而Brocadiaceae spp.略有下降,Ottowia spp.和Denitratisoma spp.急剧增加。Zhang等人(2020)报道,进水中的低浓度富里酸(25~65 mg/L)促进了厌氧氨氧化反应器的NRR,而高浓度富里酸(>80 mg/L)抑制了反应器的性能。后续的微生物群落分析表明,Ca.Jettenia和Ca.Kuenenia也能高富里酸中存活下来。据推测,AnAOB和HB之间的代谢物交换可能有助于AnAOB在不利条件下生存。相比之下,HB可能在过量有机物存在的情况下强于功能菌群,导致工艺性能恶化。据报道,在具有过量易生物降解有机物和难生物降解有机物的厌氧氨氧化系统中,异养生物的逐渐生长,HB与AnAOB之间的竞争加剧。总之,包括含量、生物降解性和组成在内的有机物特征对微生物群落施加不同的选择压力,可以重塑功能种群与HB之间的关系。我们的结果表明,在处理THP-AD废水时,功能种群和HB实现了平衡的共生并维持了稳定的脱氮性能。
通过PCoA分析进一步评估了反应器运行过程中微生物群落组成的演变。结果显示来自相同运行时期的污泥样品之间没有存在明显的聚类(图4,C)。相比之下,基于污泥粒径的样品有明显的分离(图4,D)。絮状污泥与颗粒污泥和混合污泥在PCoA 1方向上明显分离(43.2%解释度),Nitrosomonas europaea和f_Saprospiraceae对PCoA 1轴的贡献最大。来自颗粒污泥和混合污泥的样品沿PCoA 1轴紧密聚集(图4,D)并沿PCoA 2轴分离。Ca.Jettenia和Thermomicrobiales主要贡献于PCoA 2轴。这些结果表明,与絮状污泥相比,颗粒污泥与混合污泥具有更多的相似性。
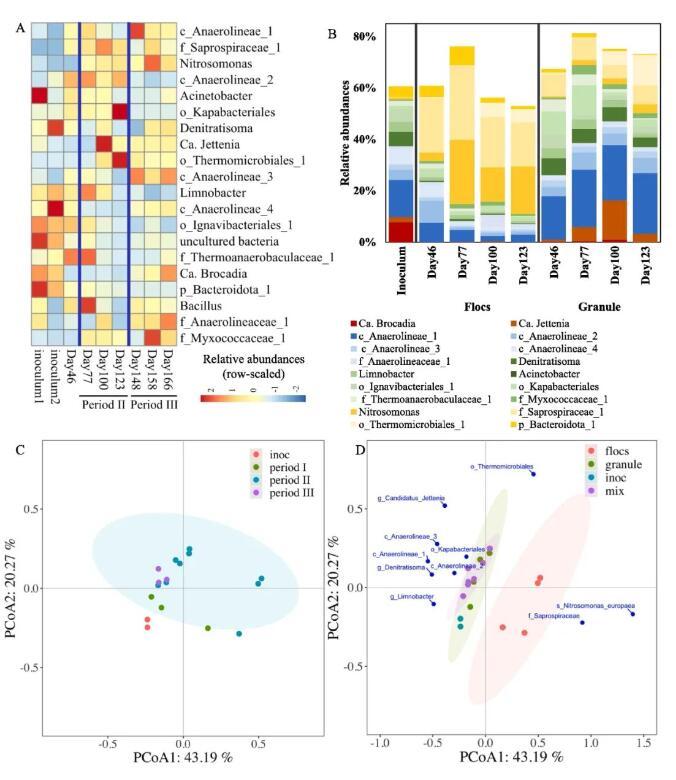
图4 PN/A反应器的微生物群落分析。A:混合污泥中微生物群落最丰富的20个属;B:絮状污泥(直径<200μm)和颗粒污泥(直径>200μm)中20个最丰富属的丰度。C:按运行期分组的微生物群落的PCoA分析。D:按颗粒大小分组的微生物群落的PCoA分析。
4 PN/A工艺的代谢潜力
微生物群落的功能由PICRUTs 2预测,并由KEGG数据库注释。根据注释结果,碳水化合物代谢、辅因子和维生素的代谢以及能量代谢是主要的预测途径。此外,翻译、膜转运、细胞生长和死亡途径丰度较高。在PN/A反应器运行期间,未检测到微生物代谢的明显变化。在微生物氮转化潜力方面,发现了负责反硝化、硝化、硝酸盐同化、异化硝酸盐还原成铵(DNRA)的基因(图5,A)。至于硝化潜力,hao基因在反应器运行过程中逐渐富集并在第100天达到峰值,这与PN/A反应器的好氧NH4+氧化能力增强一致。微生物群落显示出高反硝化潜力,群落中编码了完全反硝化途径(NO3-→NO2-→NO→N2O→N2)。编码硝酸还原酶(nar)的基因显示出高丰度,其次是编码亚硝酸还原酶(nir)、NO还原酶(nor)和N2O还原酶(nos)的基因。总体而言,运行期间反硝化潜力相对稳定。DNRA潜力不容忽视,可能有助于系统氮转化(图5,A)。
研究检测了40个最丰富的OTU的氮转化能力,其中15个OTU编码氮转化基因(图5,B)。Nitrosomonas-AOB编码amo和hao,显示完整的NH4+到NO2-途径。Ca.Jettenia-AnAOB编码hao和nar。许多编码nar的OTU,隶属于Denitratisoma、Limnobacter和f_Commonadaceae,表明它们具有将硝酸盐还原为亚硝酸盐的能力。值得注意的是,Denitratisoma编码nar、nirD和nor,能够将NO3-还原为N2O,这意味着Denitratisoma spp.是N2O生产者,这在之前的研究中也有记载,表明Denitratisoma oestradiolicum可能有助于厌氧氨氧化反应器中的N2O生产。注释为o_Thermomicrobiales_1的高度丰富OTU编码了nirK,它也可能是N2O生产者。同时,o_Ignavibacteriales、g_Ignavibacterium和f_Saprospiraceae编码nos,可能将N2O还原为N2。值得注意的是,根据PICRUTs 2的基因功能预测,丰富的异养细菌Anaerolineae没有编码任何氮转化基因。相反,之前的一项研究基于宏基因组学方法发现厌氧氨氧化颗粒污泥(本研究接种物)中的Anaerolineae编码包括nar和nir的反硝化基因。与此相同,许多研究报告称,Anaerolineae编码反硝化基因并可以进行DNRA过程。因此可见,基于16S rRNA扩增子测序的功能预测可能不完整,导致Anaerolineae spp.的氮转化基因在预测中缺失。
Denitratisoma spp.是微生物群落中丰富且潜在的重要反硝化菌,尽管其丰度在THP-AD废水运行期间略有下降。另一方面,Anaerolineae的丰度则随着反应器的运行显著增加。这种差异可能是由HB对THP-AD废水中有机物的适应引起的。在厌氧氨氧化工艺中已发现异养生物能利用细胞裂解、腐烂和可溶性微生物代谢物等来源的惰性有机物作为碳源。Denitratisoma的存在于厌氧氨氧化工艺中得到普遍鉴定,根据其栖息地的营养状况,可将其划分为兼性化学无机营养或化学有机营养细菌。
Denitratisoma能使用的电子供体包含易生物降解的有机物(如VFA)和难以生物降解的物质(如对二甲苯和雌酮),其电子受体可以是硝酸盐和氧。研究表明,两阶段PN/A工艺去除了进水中的含氧有机化合物、蛋白质和腐殖质,厌氧氨氧化反应器中反硝化菌Denitratisoma的丰度增加。因此,Denitratisoma可以通过使用可生物降解的有机物来适应RW水质。Anaerolineae spp.隶属于Chloroflexi,是能够与DNRA结合进行碳水化合物发酵的细丝细菌。Anaerolineae spp.在厌氧氨氧化工艺中表现出非凡的适应性。据报道,在处理THP-AD废水、添加葡萄糖的合成废水和生活污水的厌氧氨氧化反应器中,Anaerolineae spp.丰度增加。发酵厌氧菌主要依赖于大分子有机物生长。因此,据报道,在厌氧氨氧化工艺中发现的Chloroflexi spp.能编码和表达一系列细胞外蛋白酶、淀粉酶和细胞外肽和氨基酸的转运蛋白。因此,Anaerolineae可能具有在THP-AD废水中利用有机物生长的优越能力。HB对电子供体的偏好可以解释微生物种群的差异增长趋势。这项研究还表明Anaerolineae很好地适应了废水,原因可能是其受益于惰性有机物质。
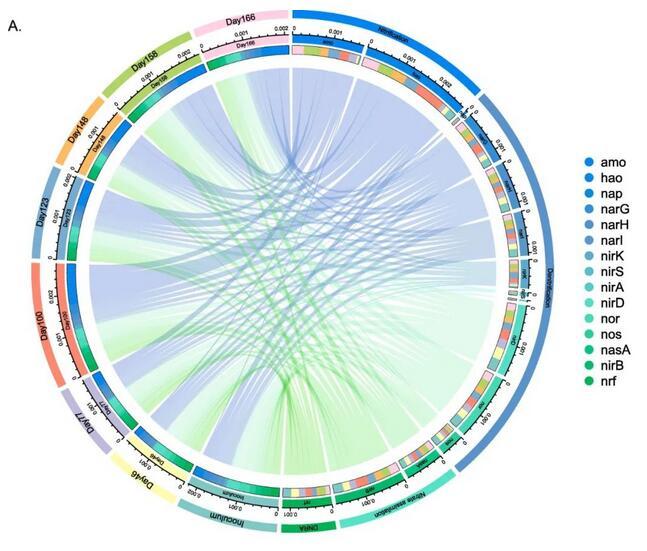

图5 PICRUSt 2预测的微生物群落功能。A:参与氮转化的基因。B:基于PICRUSt预测的由高丰度OTU编码的氮转化基因。蓝色表示OTUs编码了相应的基因,浅灰色表示OTUs没有编码相应基因。
结论
PN/A工艺在处理THP-AD废水时实现了0.58±0.06 gN/(L·d)的NRR。尽管THP-AD废水含有高COD浓度,但由于可生物降解的有机物供应有限,THP-AD废水中的异养反硝化作用可以忽略。THP-AD废水可能会影响异养N2O途径。功能微生物(AnAOB和AOB)在反应器运行期间保持活跃并不断增长。Anaerolineae在微生物群落中占主导地位,并在THP-AD废水处理期间增加,而反硝化细菌Denitratisoma略有减少。总体而言,HB和功能微生物的平衡共生确保了工艺性能的稳定性。
 相关新闻
相关新闻