热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
研究简介:介绍了卡他莫拉菌(Moraxella catarrhalis)的生物学特性及其在人类健康中的重要性,特别是它在引起呼吸道疾病方面的作用。文章强调了卡他莫拉菌从先前被认为是无害的上呼吸道共生菌,到现在被确认为能够引起上下呼吸道疾病,包括在婴幼儿中常见的急性中耳炎,以及在成人中可能导致慢性阻塞性肺病(COPD)的感染性加重。本研究重点放在了卡他莫拉菌的生物膜生长模式,这是一种在黏膜表面与其他共生细菌共同存在的形态。在生物膜中,特定的基因表达会上调,包括与反硝化途径相关的基因。反硝化是一种细菌在缺氧条件下利用氮氧化物作为电子受体进行代谢的过程。在卡他莫拉菌中,研究者发现了两个关键的开放阅读框(ORFs),aniA和norB,它们编码的蛋白质分别参与亚硝酸盐和一氧化氮的还原过程。nsrR基因编码了一个转录调控因子,对aniA和norB基因的表达具有抑制作用。当nsrR基因被失活时,aniA和norB的表达增加,这与亚硝酸盐对卡他莫拉菌生长的抑制有关。
研究还发现,nsrR基因的突变体在亚硝酸盐存在下的生长受到抑制,但这种抑制可以通过aniA基因的突变或nsrR基因的互补来逆转。增进了我们对卡他莫拉菌如何在宿主中生存和致病的理解,而且可能为开发新的治疗策略提供了潜在的靶点。通过了解这些基因和它们在细菌适应和致病过程中的作用,我们可以更好地控制和预防由卡他莫拉菌引起的感染。
Unisense微电极分析系统的应用
Unisense微电极分析系统被用于定量测量卡他莫拉菌(Moraxella catarrhalis)在不同条件下对氮氧化物的代谢活动。首先将卡他莫拉菌的野生型和突变型菌株在BHI肉汤中培养至OD600(光学密度在600纳米处)达到2.0,然后洗涤并重新悬浮在新鲜的BHI肉汤中至OD600为1.0。为了研究亚硝酸盐(NO2-)的代谢,向细胞悬浮液中添加最终浓度为5 mM的NaNO2。使用氧气不敏感的、N2O特异性的探针(N2O-50-3112)连接到皮安表(PA2000),并通过模数转换器(ADC 216)记录数据。在添加NaNO2后,监测细胞悬浮液中N2O的产生。收集不同时间点的N2O的浓度数据,以评估卡他莫拉菌对这些氮氧化物的代谢速率和途径。
实验结果
研究确定了NsrR是一个关键的转录调控因子,它在卡他莫拉菌中调控着一个截断的反硝化途径。NsrR通过抑制aniA和norB两个基因的表达来调控亚硝酸盐还原酶和一氧化氮还原酶的活性。在nsrR基因失活的情况下,aniA和norB基因的表达显著上调,这表明NsrR对这些基因具有负调控作用。这种调控对卡他莫拉菌在不同环境条件下的生存和适应至关重要。nsrR基因的突变体在低水平的亚硝酸盐存在下生长受到抑制,这种抑制可以通过aniA基因的突变或nsrR基因的互补来逆转。这表明NsrR在调节卡他莫拉菌对亚硝酸盐的敏感性方面发挥着重要作用。通过使用微电极技术和分子生物学方法,揭示了卡他莫拉菌能够通过AniA和NorB蛋白分别还原亚硝酸盐和一氧化氮,并且能够将一氧化氮进一步还原为一氧化二氮。卡他莫拉菌可能利用其截断的反硝化途径在氧限制条件下更有效地呼吸,并保护自己免受宿主防御机制产生的一氧化氮的侵害。这项研究增进了我们对卡他莫拉菌如何在宿主中生存、适应并引起疾病的理解,为开发新的防治策略提供了可能的靶点。

图1、参与截短脱氮途径的卡他莫拉菌基因产物示意图及相关突变体的构建。(A)卡他莫拉菌的截短脱氮途径包括所示的前三个酶解步骤。卡他莫拉菌显然缺乏将N2O还原为N2的能力(用括号表示)。(B至D)包含norB、nsrR和aniA基因的卡他莫拉菌染色体位点示意图,(B)野生型O35E菌株、(C)O35E nsrR突变体和(D)O35E aniA突变体的侧翼区域。箭头表示用于PCR的不同引物的相对位置。
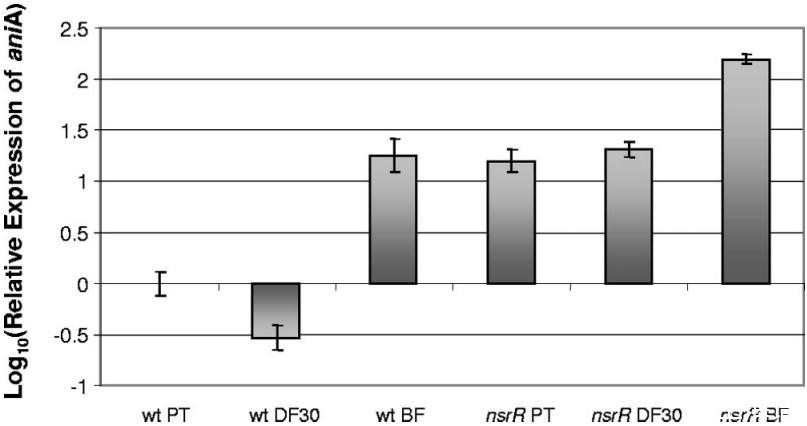
图2通过qRTPCR测定卡他莫拉菌ETSU-9野生型和突变株在不同生长条件下aniA的表达。从野生型ETSU-9(wt)和ETSU-9 nsrR突变体(nsrR)细胞中分离出总RNA,这些细胞在含30μM Desferal(DF30)的肉汤和连续流生物膜系统(BF)中生长,在铁限制条件下处于浮游状态(PT)。qRTPCR如“材料与方法”中所述进行。
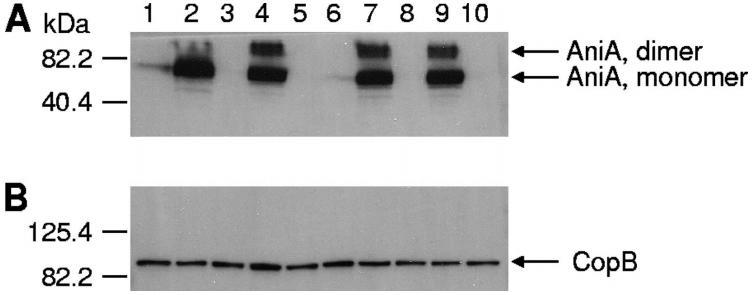
图3、NsrR对野生型、突变型和互补突变型卡他莫拉菌AniA蛋白表达的影响。用多克隆AniA抗血清(A)或CopB特异性单克隆抗体10F3(B)作为一抗,对菌株的全细胞裂解液进行Western印迹分析。泳道1,野生型菌株O35E;泳道2,O35E nsrR突变体;泳道3,O35E aniA突变体;泳道4,O35E nsrR(pWW115);泳道5,O35E nsrR(pWW150);第6道,野生型ETSU-9;第7道,ETSU-9 nsrR突变体;第8道,ETSU-9 aniA突变体;第9道,ETSU-9 nsrR(pWW115);第10道,ETSU-9 nsrR(pWW150)。推测的AniA单体和二聚体的位置用A组右侧的箭头表示。CopB外膜蛋白被用作载荷对照,其位置在B组右侧用箭头表示。分子质量标记物的位置在各图的左侧。
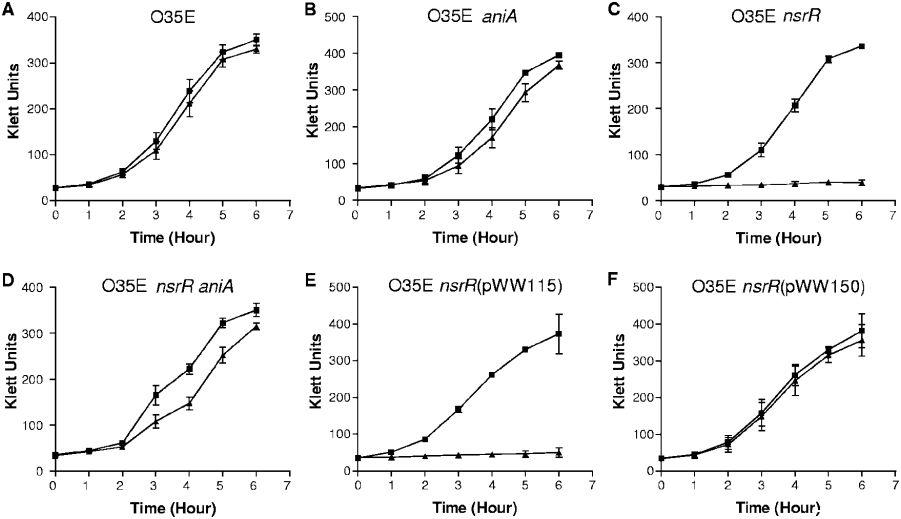
图4、NO2-对卡他莫拉菌野生型、突变型和互补突变株生长的影响。野生型菌株O35E(A)、O35E aniA突变体(B)、O35E nsrR突变体(C)、O35E nsrR aniA突变体(D)、O35E nsrR(pWW115)(E)和O35E nsrR(pWW150)(F)的细胞在BHI培养基(黑正方形)或含5mM NaNO2的BHI培养基(黑三角形)中生长。大部分数据为三个独立生长实验的平均值;B组数据例外,为两个独立实验的平均值。
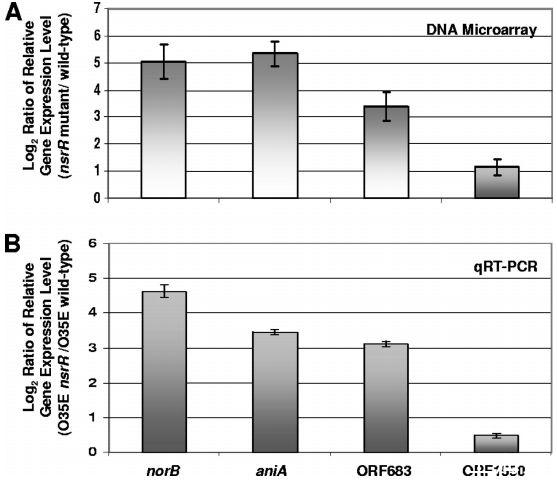
图5、确定受NsrR调控的卡他莫拉菌基因。(A)按照材料与方法中的描述,对从野生型菌株O35E、7169和ETSU-9以及这些菌株的nsrR突变体中分离的总RNA进行了DNA微阵列分析。图中显示了在所有三个nsrR突变体中持续上调至少两倍(P<0.002)的基因数据。误差条表示标准偏差。(B)qRT-PCR分析norB、aniA、MC ORF 683和MC ORF 1550在菌株O35E的nsrR突变体和野生型细胞中的相对表达水平。qRT-PCR测量的内源对照是MC ORF 1234的表达。最大和最小相对表达水平用误差条表示。
结论与展望
卡他莫拉菌在生物膜中的生长导致两个开放阅读框(ORF)aniA和norB的明显上调,这两个开放阅读框被预测为分别编码亚硝酸盐还原酶和一氧化氮还原酶。研究表明,位于aniA和norB之间的一个ORF被命名为nsrR,它编码一个预测的转录调节因子。通过DNA微阵列分析和定量反转录酶PCR测定,nsrR失活导致三种不同的卡他莫拉菌菌株中aniA和norB的表达增加。在nsrR突变体中转入野生型nsrR基因会导致AniA蛋白表达量减少。DNA微阵列分析表明,另外两个ORFs(MC ORF 683和MC ORF 1550)在nsrR突变体中也持续上调。与野生型细胞相比,nsrR突变体细胞消耗亚硝酸盐和一氧化氮的速度更快。然而,低浓度的亚硝酸钠完全抑制了nsrR突变体的生长。在nsrR突变体中引入aniA突变后,亚硝酸盐对生长的抑制作用明显逆转,而野生型nsrR基因的反式存在则完全逆转了这种抑制作用。NsrR对aniA表达的调控对亚硝酸盐敏感,而NsrR对norB的调控对一氧化氮敏感。本研究提供了对卡他莫拉菌反硝化途径及其调控机制的深入见解,并揭示了这些机制在细菌致病性中的潜在作用。unisense微电极技术在本研究的应用对于理解卡他莫拉菌在生物膜状态下的代谢途径,以及NsrR调控蛋白如何影响这些代谢途径至关重要。通过这些数据,研究人员能够更深入地了解卡他莫拉菌如何在宿主中生存,以及它如何响应和利用环境中的氮化合物。这对于揭示细菌的致病机制和开发新的治疗策略具有重要意义。
