热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
3.3 CHT对反硝化酶、基因丰度和微生物群落结构的影响
为了确定CHT对反硝化酶活性的影响,评估了四种反硝化还原酶(NAR、NIR、NOR和NOS)的活性(Saggar等人,2013)。如图3a和b所示,对照处理和CHT处理在NAR活性方面没有显着差异。对照组的NIR活性显着高于C25处理组(P<0.01)。这与NO2的变化是一致的−在本研究中,这表明CHT显着抑制NO2−还原过程,产生NO2−高浓度CHT的累积。与对照组相比,C25处理组的NOR活性显着降低29.4%(P<0.05),而C5和C10处理组的NOR活性无显着差异。此外,高CHT添加处理(C10、C25)的NOS活性也显着降低(P b 0.01),C25处理的NOS活性比对照(C0)下降80.3%。从结果来看,NOS活性的下降大于NOR活性的下降,这表明C25处理中NOS活性的显着抑制可能导致CHT施用后N2O排放量增加。郑等人(2014b)还发现,大量N2O排放与相应的反硝化还原酶活性抑制有关。如上所述,CHT对NIR、NOR和NOS活性的显着抑制可能是CHT对反硝化过程产生负面影响的原因。
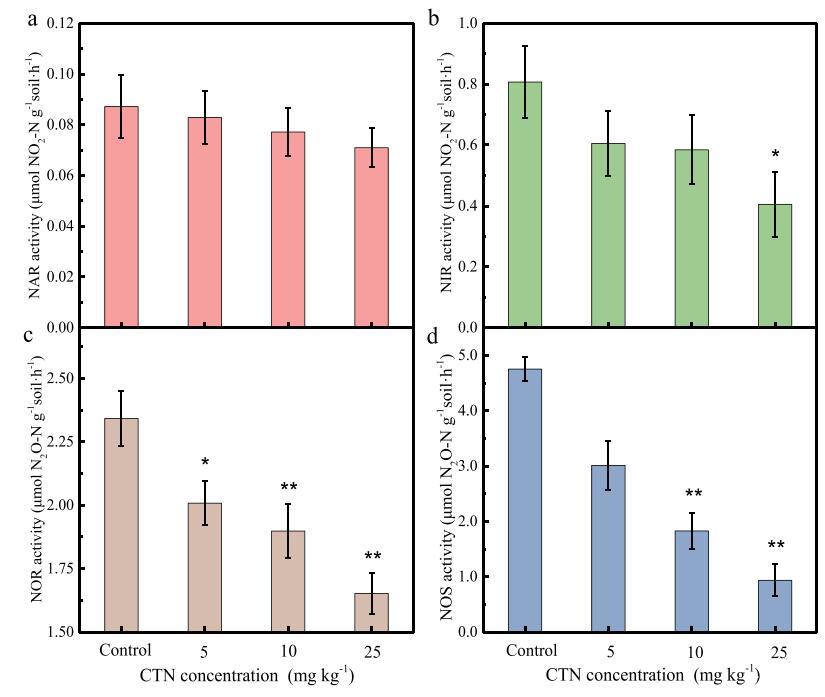
图3.百菌清(CHT)对反硝化过程中NAR(a)、NIR(b)、NOR(c)和NOS(d)活性的影响。数据显示为三次独立测量的平均±标准偏差。Tukey t检验,与对照组相比,*P b 0.05,**P b 0.01。
作为反硝化基因表达的产物,反硝化还原酶活性与反硝化基因丰度相关。由于CHT对反硝化的抑制是一个剂量反应过程,因此NO3存在显着差异−C0和C25处理之间的减少和N2O排放。因此,本研究选择CHT对C0和C25处理下反硝化基因丰度的影响进行研究。如图S3所示,功能基因narG、nirS、nirK、norB和nosZ的拷贝数在104到107拷贝之间−1干燥土壤。对照组或CHT处理中narG基因的拷贝数高于其他四个基因的拷贝数。这一结果与之前的研究一致,之前的研究报告表明,narG基因在大多数自然土壤中的丰度最高(李等,2017;张等,2016)。对照处理的nirS、nirK和norB基因丰度分别为5.06×104、4.20×104和4.65×104−1干土。与之前研究中报告的基因丰度相比(Henry等人,2006),本研究中的nirS、nirK和norB基因丰度明显较低。这可能与实验土壤中反硝化细菌丰度的差异有关。值得注意的是,本研究中的nirS基因拷贝数低于nirK基因拷贝数,这与之前的研究不一致(Shrewsbury等人,2016)。nirS和nirK基因之间丰度的差异可能取决于土壤性质,如pH值、养分和土壤铜(Cu)含量(Enwall等人,2010)。我们的结果表明,本研究中的茶园土壤比nirS基因更适合nirK基因,并且可以促进nirK基因丰度的增加。此外,thenosZ基因的拷贝数(约105个拷贝)为−1干土)与之前研究(Henry等人,2006)中测量的其他自然土壤相似,比nirK、nirS和norB基因的拷贝数大一个数量级。
图4显示,CHT添加降低了反硝化菌中的基因丰度,但程度不同。方差分析显示,对照组之间的差异不显着(1.81×107μg)−1干土)和C25处理(1.76×107 g−1干土)的narG基因数据。然而,与对照处理相比,CHT处理的nirK、nirS和norB基因丰度分别下降了31.6%、22.1%和12.7%。CHT处理中nirK(P b 0.05)和nirS(P b 0.05)基因丰度的显着降低可能导致反硝化菌中NIR活性的降低,这可能是NO2累积的另一个原因−.C25处理的nosZ基因拷贝数为2.47×105拷贝−1干土,与对照处理(5.01×105拷贝)相比减少了50.7%−1干土)(P b 0.01)。nosZ基因丰度的降低可能导致NOS活性的降低,这将显着抑制从N2O到N2的减少。nosZ基因丰度的降低(50.7%)远高于norB基因丰度的降低(12.7%),这表明CHT对N2O消耗的抑制作用比N2O产生的抑制作用强得多,导致N2O排放量显着增加。
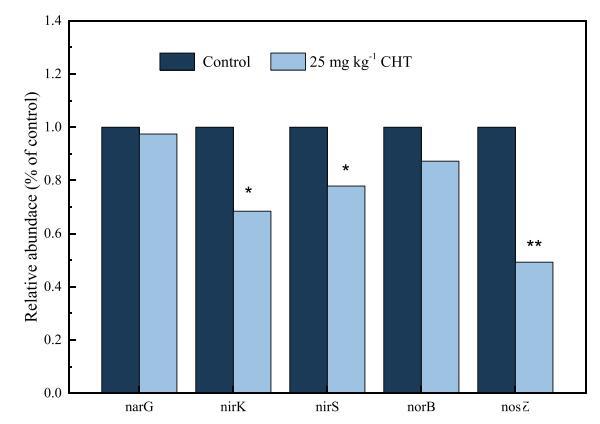
图4.百菌清(CHT)对对照组和25 mg kg体重组narG、nirS、nirK、norB和nosZ的相对基因丰度的影响−1 CHT处理。Tukey t检验,与对照组相比,*P b 0.05,**P b 0.01。
从对照和CHT处理中获得的细菌在门和类群水平上的分类学如图S4所示。厚壁菌、放线菌、氯屈曲菌和变形菌在两种处理中占优势。与对照处理相比,CHT处理后厚壁菌和放线菌的相对丰度分别从81.6%和9.4%下降到80.9%和5.2%。在类别水平上,在C0和C25处理中均检测到α、β、γ和δ-变形菌,在之前的研究中,据报告这些变形菌与氮循环过程有关(王等人,2016)。从C0到C25处理,α、β和δ变形菌的丰度分别下降了2.3%、0.04%和0.1%。两种处理中检测到的前25个属如图S5所示。芽孢杆菌作为自然系统中潜在反硝化的贡献者和nosZ基因的持有者(Verbaendert等人,2011),在CHT处理后从9.8%下降到8%。如上所述,对照和C25处理之间的微生物群落没有明显变化。这表明CHT的应用可能不会通过改变微生物群落来影响反硝化过程,但可能会影响微生物的细胞内代谢。测定ETSA值和ATP含量以评估CHT对微生物代谢过程的影响。
3.4 CHT对反硝化相关代谢过程的影响
反硝化菌接受来自有机物代谢过程和电子传输链的电子,以完成反硝化过程(Chen和Strous,2013)。碳源代谢和电子传递过程与反硝化过程密切相关。因此,通过测量ETSA值和ATP含量来确定CHT对这两个过程的影响。如图5a所示,ETSA值为0.186μgO2·mg−1蛋白质·min−C0为1,C5、C10和C25分别下降了34.9%、43.5%和46.7%,这表明较高的CHT应用可以大大降低ETSA值。对照组和低CHT浓度(C5)之间的ETSA值没有显着差异。同时,高浓度CHT(C10(P b 0.05)、C25(P b 0.05))显着抑制电子传递系统。此前的一项研究还报告称,ETSA值的降低与反硝化过程的抑制有关(Wan等人,2016年)。C25处理中ETSA值最低的记录表明CHT对电子传输过程有抑制作用,这将导致反硝化过程中可用的电子更少。
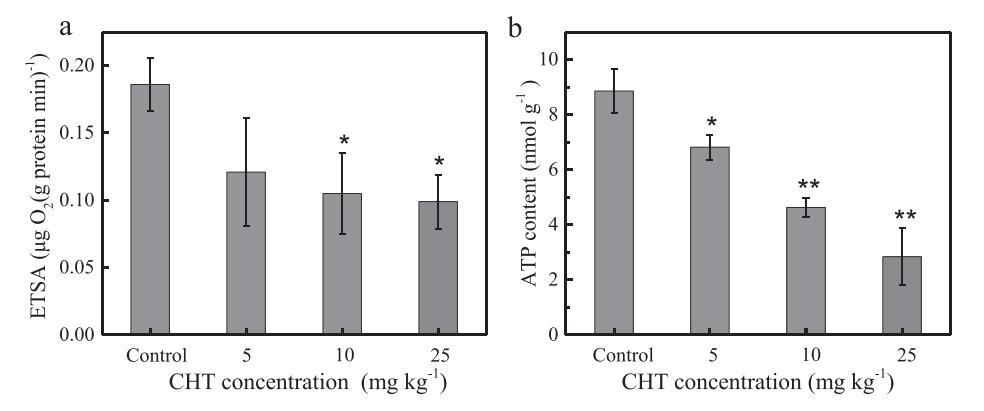
图5.对照组和百菌清(CHT)处理(5、10和25 mg kg)中电子传递系统活性(ETSA)(a)、三磷酸腺苷(ATP)含量(b)的值−1)72小时后,误差条表示三次试验的标准偏差。Tukey t检验,与对照组相比,*P b 0.05,**P b 0.01。
图5b表明,土壤中的ATP含量将随着CHT浓度的增加而降低。ATP浓度为8.86 nmol g−1,但显着下降至4.63和2.85 nmol g−分别在C10(P b 0.05)和C25(P b 0.01)处理中为1。ATP被广泛认为是参与细胞内能量转移的货币单位,可以在碳源代谢和电子传递链中产生(Knowles,1980)。Hammes等人(2010年)报告说,ATP可能是微生物生存能力的有用指标。在本研究中,对照治疗中的ATP浓度与之前研究中报告的相似(邱等人,2016)。对于C25处理,ATP浓度的降低可能表明CHT对微生物活性具有抑制作用,例如碳源代谢以及电子传输和消耗过程(Junge和Nelson,2015)。结合ETSA值和ATP含量的结果,CHT应用将抑制电子传输过程。因此,到脱氮过程的有限电子传输可能是CHT应用后抑制脱氮过程的一个因素。
3.5土壤反硝化作用与分子指标的关系
尽管3号−还原过程和相关还原酶活性已由之前的研究确定,我们对NO3之间关系的理解−去除过程和还原酶活性仍不清楚。因此,建立了线性关系,将反硝化过程与分子指标联系起来,以便更好地在分子水平上理解反硝化过程。
通过相关分析建立了反硝化作用与分子指标之间的关系(表S3)。一些关键相关性如图6所示。如图6a所示,NO3−-氮去除率与NAR活性呈正相关,因为NAR可以减少NO3−至NO2−.最终NO2-N浓度与NAR和NIR活性呈负相关(图6b),NIR和最终NO2的拟合线斜率−-氮浓度大于NAR和最终NO2的浓度−-氮浓度。这种关系表明CHT对NO2的抑制作用−还原过程大于NO2上的还原过程−导致NO2累积的生产过程−.同样,最终N2O浓度与NOR和NOS活性呈负相关(图6c)。根据相关分析数据(表S3),NOS的相关系数(−0.968)高于NOR(−0.920)与N2O浓度相关。这表明NOS活性对CHT比NOR活性更敏感。因此,NOS活性的降低是导致N2O积累的关键因素。至于微生物活性指标,图6d和e表明,ATP含量与四种反硝化酶的活性呈正相关,表明反硝化过程的抑制可能是由于微生物代谢的下降。ETSA值与NOR和NOS活性显着正相关。这些结果表明,电子传递的减少可能是CHT对反硝化产生急性抑制作用的另一个原因。
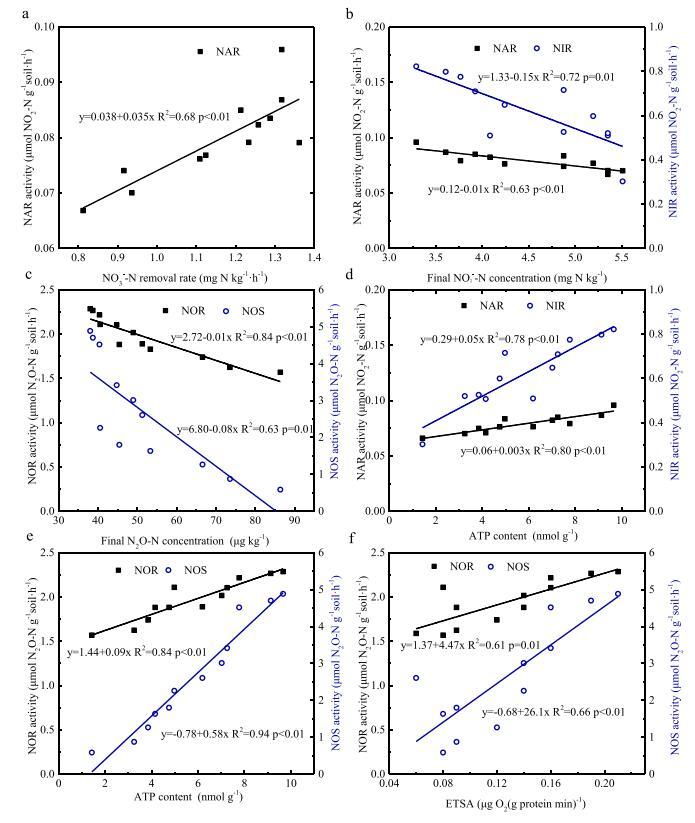
图6.反硝化酶活性与NO3之间的线性关系−-N去除率(a);NO2号−-氮浓度(b);N2O-N浓度(c);三磷酸腺苷(ATP)含量(d)(e);和电子传输系统活性(ETSA)值(f)。
3.6环境意义
近几十年来,全球茶园面积显着增长,从2002年(265万公顷)到2014年(437万公顷),增长了64.9%。茶园区域的反硝化过程令人担忧,因为它会产生温室气体N2O。我们的结果表明,将CHT的施用量从0增加到25 mg kg−1抑制NO3−去除效率从83.8%提高到54.1%,但N2O排放量增加了94.8%。当533.4 kgN hm时−在茶园区域施用2%的氮肥和5 mg kg−大田施用CHT 1,年N2O-N排放量增加3.0×106kg,年未还原NO3量增加−-氮保持在5.48×108 kg。因此,CHT在茶园土壤上的应用将对全球气候产生负面影响,导致更多的NO3−从土壤中渗入水中,导致富营养化加剧。虽然施用CHT会增加茶园土壤反硝化过程中的N2O排放,但也应考虑通过CHT控制茶树病害。今后,应严格限制农药的施用量,以平衡害虫控制和环境风险。
4、结论
在72小时的实验室试验中,研究了CHT对茶园土壤的急性影响。结果表明,CHT的应用导致了NO3的减少−去除效率、NO2积累−,以及N2O排放量的增加。CHT的施用抑制了反硝化酶的活性,降低了反硝化基因的丰度。此外,本研究表明,CHT应用导致ETSA和ATP含量降低。需要进一步研究以确定CHT对土壤反硝化的慢性影响及其代谢产物对土壤健康的潜在毒性。
致谢
本研究得到了中央高校基础研究基金(编号:106112017CDJXY210005、106112017CDJQJ218843)的资助。我们还感谢弗雷德里克·库隆教授和匿名评论员对本手稿的早期草稿提出的建设性建议和意见。
