热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
C3植物是最重要的光合生物之一,在光照条件下,其类囊体膜上光驱的电子流与基质内质子的跨膜转运相耦合。质子在囊腔内的积累产生了跨膜质子动势(pmf),其驱动ATP合成酶的构象发生转变并合成ATP分子。质子动势的一个重要组分,即跨膜电势差(ΔΨ),是反映原初光合能量转化的一个重要指标量,它会使色素分子(包括叶绿素分子以及胡萝卜素分子等)对光能的最大吸收峰迁至515 nm处。515 nm最大吸收峰的形成除了有ΔΨ的贡献,也有来自“光散射”效应的叠加。因此,对于ΔΨ的实验测量一直是一个难题,并且在国内,关于ΔΨ的实验测量鲜见报道。以烟草叶作为样品,使用双通道可变振幅式叶绿素荧光仪,并搭配最新设计的P515/535模块,测量样本在单周转饱和光(ST)、1 s可见光、60 s可见光的P515光谱信号变化情况。实验结果可为研究C 3作物高光效品种选育提供技术支持。
光合作用是地球生物圈最重要的生化反应过程之一。它能够固定大气中的CO2生成有机物,为人类的生存提供物质基础。C3植物是最为重要的放氧型光合生物之一,其光合作用的原初阶段发生在光系统反应中心,集光复合体吸收太阳光能,通过激子传递机制,直至被光反应中心色素分子捕获,用来催化光反应中心的电荷分离。除了用于光合作用外,光能也能以叶绿素荧光辐射的形式而损失。随后,水分子在光放氧复合体中裂解,释放出氧气、质子和电子。释放的电子经线性电子传递链传递至铁氧还蛋白-NADP+-氧化还原酶(FNR),还原NADP+并合成NADPH。同时,光合反应的电子传递过程和质子跨类囊体膜的转运相偶联,形成跨膜的质子梯度。根据Mitchell的化学渗透偶联假说,质子梯度和跨膜电势差在能量上是等价的,并且共同作用形成了质子动势(pmf)。
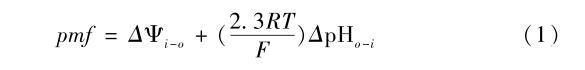
其中i和o代表类囊体内腔和外周基质,ΔΨ和ΔpH为电势差和pH值差,R、T和F有其通常的物理意义。
质子动势为ATP合成提供能量来源,最终,合成的ATP和NADPH被用于Calvin-Benson循环来固定CO2合成有机物。
跨膜电势在光合能量转化中扮演相当重要的角色。它不仅是光合能量传导的一个重要指针,同时,研究电势信号曲线的衰退可以得到关于类囊体膜的渗透性及其跨膜离子流强度的信。更为重要的是,跨膜电势差对电子传递链中电荷分离的稳定性以及调控电荷复合具有重要作用。跨膜电势差的测量可以使用微电极直接刺入叶绿体类囊体中,或者用膜片钳夹住叶绿体再用微电极刺入类囊体测量跨膜电压或者电流。但是上述这两种方法都会对样本产生刺入性损伤,造成电势无法预估的损失。除了微电极刺入,跨膜电势差还有一种快速、灵敏以及无损样本的探测方式,即测量P515信号。P515信号的产生是因为跨膜电势差迫使叶绿素分子,主要是胡萝卜素以及叶绿素b对入射光的最大吸收峰迁移至515 nm。本研究以烟草叶片(Nicotianatabacum)作为待测样品,使用Dual-PAM-100叶绿素荧光仪的P515/535双组件模块,测量烟草叶片在不同光信号条件下的P515信号变化情况。实验结果可为研究C3作物高光效品种选育提供技术支持。
1材料与方法
研究在珍珠岩基质中种植烟草苗(Nicotianatabacum),并施以Knop营养液,放置在Sanyo植物生长箱中培养2周,培养箱中模拟白昼黑夜时间设置为8 h/16 h,白昼时光强度为90μmol/m2·s。白昼黑夜温度分别设置为22℃/15℃。使用Dual-PAM-100叶绿素荧光仪的P515/535模块来测量P515信号。P515/535模块包含发射单元DUAL-EP515和探测单元DUAL-DP515。DUAL-EP515可以由一个支撑臂和DUAL-DP515串联连接起来。P515信号是测量叶片透过光的550 nm和515 nm的差值,即ΔA550-515 nm。选择550 nm是为了能消除“光散射”效应所产生的光吸收峰的叠加,因为535 nm反映了“光散射”效应所产生的光吸收峰迁移,535 nm的Gauss吸收峰在515 nm和550 nm所在的部分均匀一致,所以计算叶片透过光的550 nm和515 nm的差值,在理论上可以消除“光散射”效应所产生的在515 nm吸收峰的叠加。
 相关新闻
相关新闻