热线:021-56056830,66110819
手机:13564362870
热线:021-56056830,66110819
手机:13564362870
研究简介:研究表明,绝经前女性的心脏病发病率低于同龄男性,但这种性别优势在女性绝经后消失,这提示内源性雌激素可能具有心脏保护作用。雌激素已被证实具有多种生物学效应,其中抗氧化应激是与心脏保护相关的关键效应之一。雌激素能够减少心肌中的活性氧种类生成,并通过抑制缺血/再氧合诱导的氧化应激来减少细胞凋亡。硫化氢(H2S)作为一种新发现的气体信号分子,硫化氢在心肌中主要由CSE合成,并且已被证明具有心脏保护作用,其机制之一是抗氧化应激。之前的研究表明,雌激素治疗能够改善氧化应激,恢复因去卵巢手术导致的大鼠心肌中CSE表达和硫化氢生成的降低,提示雌激素可能通过维持心肌中CSE的表达来保护心肌细胞免受氧化应激的损害。
本研究旨在证实CSE有助于17β-雌二醇(E2)对心肌细胞氧化应激的心脏保护作用,并阐明亚型雌激素受体在E2对CSE的调控中的作用。由于Sp-1与多种细胞中CSE的调控有关,而已知雌激素可调控心肌中的多种miRs,因此研究人员进一步确定Sp-1是否介导了雌激素对CSE的调控,并确定心肌中调控CSE的miRs。
Unisense微呼吸系统的应用
使用微型硫化氢微呼吸瓶(型号H2S-MRCh;Unisense)和Unisense PA2000放大器测量了原代培养的大鼠新生心肌细胞产生硫化氢的实时动力学。收集心肌细胞并在不含FBS的培养基中清洗,培养基中用20mM HEPES代替碳酸氢钠,以防止封闭的微呼吸瓶腔室(Unisense)中形成二氧化碳气泡。最终细胞团以1×107个细胞/毫升的浓度重悬,并将0.1毫升细胞悬液注入呼吸瓶。此外,为避免硫化氢自发氧化,在加入心肌细胞之前,用氮气对呼吸室中的培养基进行脱氧。微电极信号稳定后,加入L-半胱氨酸(1mmol/L,CSE的底物)和5-磷酸吡哆醛(1mmol/L,CSE的辅助因子)以刺激硫化氢生成。在添加底物和辅助因子后,在每个迹线的最初最陡斜坡处测定硫化氢生成率。细胞活力通过胰蓝排除法进行评估,在整个实验过程中,细胞活力保持在90%以上。每次实验后,根据制造商的手册,使用与实验相同的缓冲液和条件,用新鲜制备的缺氧硫化钠原液(0-100μmol/L)校准硫化氢微电极。
实验结果
研究发现雌激素17β-雌二醇(E2)通过与雌激素受体α(ERα)结合,在体外培养的心肌细胞和雌性小鼠体内上调了CSE的表达。E2能够降低心肌细胞中miR-22的表达。miR-22是一种微小RNA,能够靶向ERα和Sp-1,调控它们的表达水平。E2通过降低miR-22的表达,间接促进了特异性蛋白-1(Sp-1)的表达。Sp-1是一种转录因子,能够结合到CSE基因启动子区域的保守位点上,从而促进CSE基因的转录和表达。由于Sp-1的上调,CSE基因的表达得到了增强,导致心肌细胞中硫化氢的产生增加。硫化氢的增加有助于提高心肌细胞的抗氧化防御能力,从而保护心肌细胞免受氧化应激的损伤。E2通过上述机制,增强了心肌细胞对H2O2和缺氧/再氧合(H/R)诱导的损伤的抵抗力,显示出心脏保护作用。揭示了雌激素在性别差异心脏病发病率中可能发挥的分子机制,尤其是在绝经前女性中观察到的较低心脏病发病率。
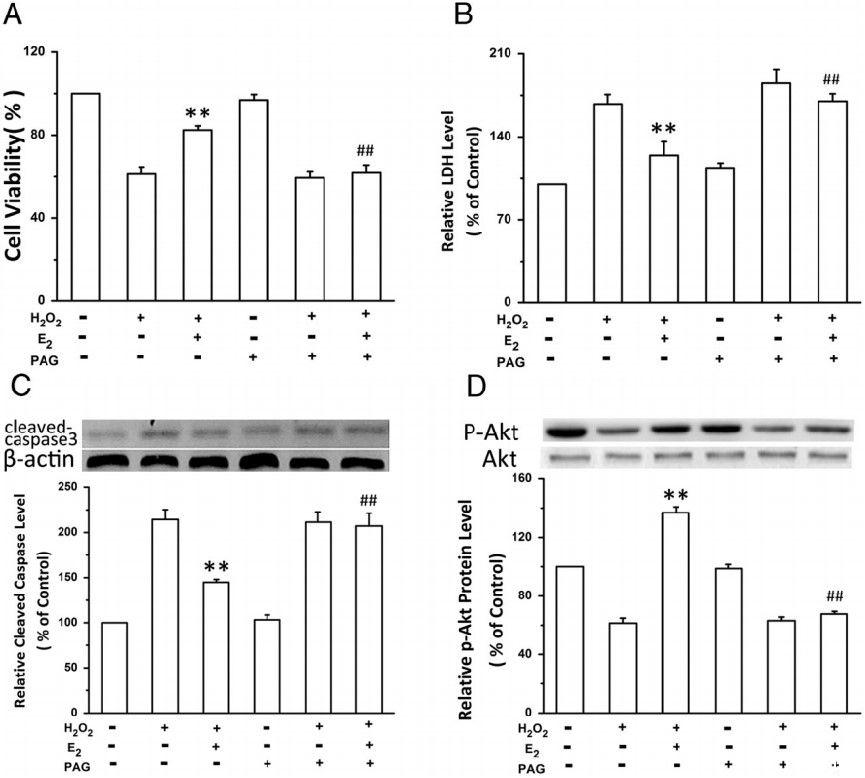
图1、CSE抑制剂减弱E2介导的原代培养新生大鼠心肌细胞氧化应激保护作用。在无CSE抑制剂(PAG,1mmol/L)或有CSE抑制剂存在的情况下,用E2(10nmol/L)培养细胞24小时。然后将细胞暴露于H2O2(200μmol/L)中4小时。通过MTT(A)和上清液LDH浓度(B)评估细胞的整体存活率。通过Western blot分析确定裂解的Caspase-3(C)和磷酸化的Akt(D)水平。
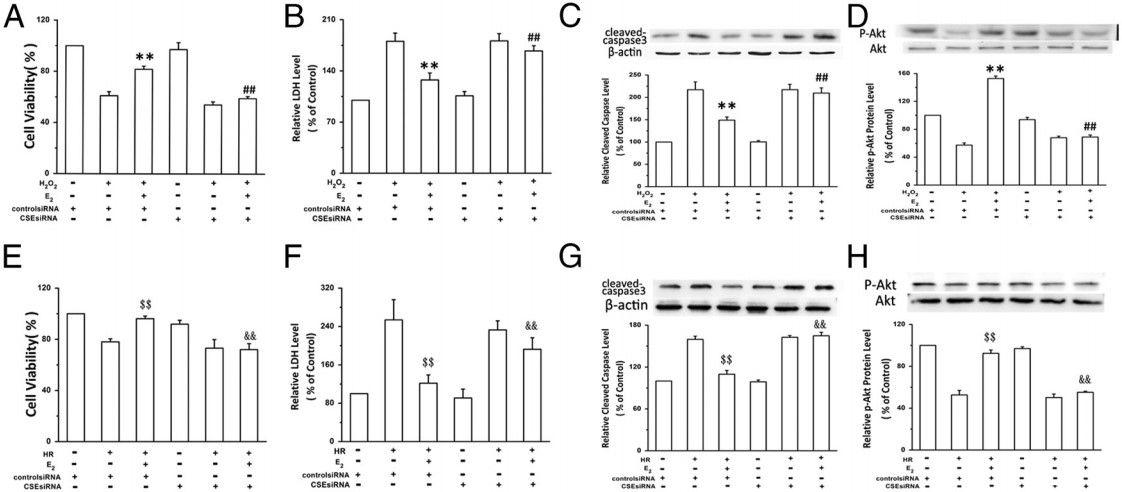
图2、CSE siRNA可减弱E2-介导的原代培养新生大鼠心肌细胞氧化应激保护作用。用对照siRNA或CSE siRNA预处理细胞24小时,然后用或不用E2(10nmol/L)孵育24小时。A-D,然后将细胞暴露于H2O2(200μmol/L)4小时。E-H,然后将细胞放入厌氧室,用92%N2、5%CO2和3%O2净化24小时,然后复氧2小时。通过MTT(图A和E)和上清液LDH浓度(图B和F)评估整体细胞活力。通过Western印迹法测定裂解的Caspase-3(图C和图G)和磷酸化的Akt(图D和图H)水平。
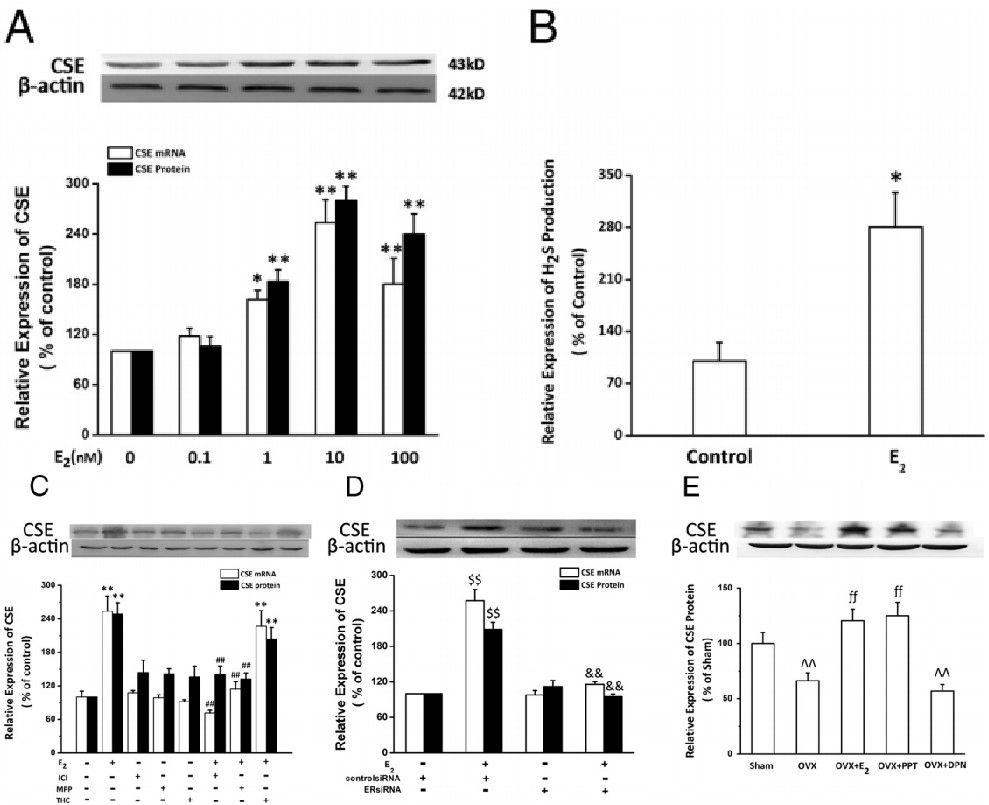
图3、E2在原代培养的新生心肌细胞中介导E2对CSE表达的调节。A,用指定剂量的E2刺激细胞24小时。采用定量实时RT-PCR和Western印迹分析分别测定心肌细胞(n=4)中CSE mRNA和蛋白的表达。B,用E2(10nmol/L)刺激细胞24小时。实时硫化氢生成测量用于确定硫化氢生成率,如材料和方法中所述(n=3)。C,在无或有非特异性ER拮抗剂ICI 182780(1μmol/L)、ERα特异性拮抗剂MPP(1μmol/L)或ERβ特异性拮抗剂THC(1μmol/L)的情况下,用E2(10nmol/L)处理细胞24小时。定量实时RT-PCR和Western印迹分析分别用于测定心肌细胞(n=4)中CSE mRNA表达和蛋白表达。D,用对照siRNA或ERαsiRNA转染细胞24小时,然后用E2(10nmol/L)处理24小时。采用实时RT-PCR定量分析和Western印迹分析分别测定心肌细胞中CSE mRNA表达和蛋白表达(n=4)。E,对雌性小鼠进行双侧卵巢切除术以清除内源性雌激素。外源性E2和ERα选择性激动剂(PPT)和ERβ选择性激动剂(DPN)均持续给药4周。采用Western印迹分析测定心肌中CSE蛋白的表达(每组8人)。
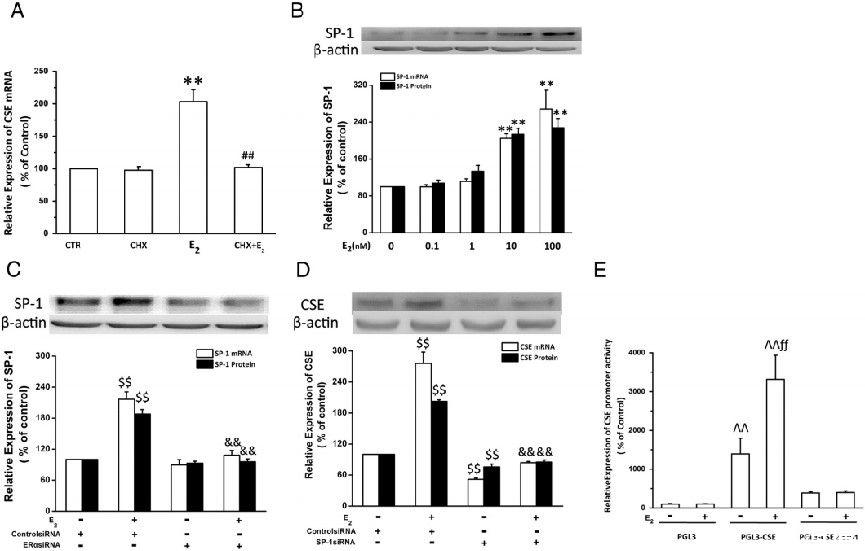
图4、Sp-1在大鼠心肌细胞中介导E2对CSE的上调。A,蛋白合成抑制剂(CHX)对E2诱导的CSE mRNA表达的影响。用CHX(10μmol/L)预处理原代培养的新生心肌细胞1小时,然后用E2(10nmol/L)刺激24小时。通过实时RT-PCR定量评估CSE mRNA的稳态水平。B、E2对Sp-1表达的影响。用指定剂量的E2处理原代培养的新生儿心肌细胞24小时。定量实时RT-PCR和Western印迹分析分别用于检测Sp-1mRNA表达和蛋白表达。C,ERα介导E2诱导的Sp-1表达。用对照siRNA或ERαsiRNA转染原代培养的新生儿心肌细胞24小时,然后用E2(10nmol/L)处理24小时。D,Sp-1在E2上调CSE表达中的作用。用对照siRNA或Sp-1 siRNA转染原代培养的新生儿心肌细胞,然后用E2(10nmol/L)处理24小时。E,E2对大鼠CSE启动子活性的影响。用大鼠CSE启动子-荧光素酶报告构建体pGL3-CSE或突变构建体pGL3-CSE SP1转染H9c2细胞,然后暴露于E2 24小时。使用双荧光素酶报告实验分析启动子活性。实时定量RT-PCR和Western印迹分析分别用于测定CSE或Sp-1 mRNA表达和心肌细胞蛋白表达(n=4)。
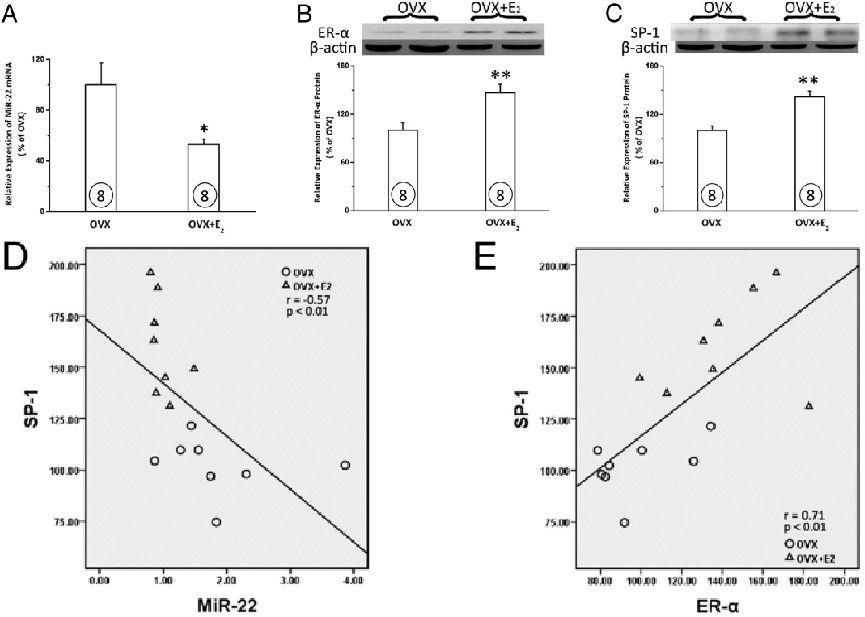
图5、E2疗法对卵巢切除大鼠心肌中miR22、ERα和Sp-1表达的影响。对雌性大鼠进行双侧卵巢切除术,以消耗内源性雌激素。外源性E2给药8周。定量实时RT-PCR和Western印迹分析分别用于测定miR-22的表达(A)以及ERα和Sp-1的蛋白表达(B)。条形图内的圆圈表示每组的人数。所有条形图均代表平均±SEM,代表性的Western印迹条带位于相应图的顶部。D,用线性回归法分析miR-22与Sp-1水平的相关性。E,线性回归法分析ERα与Sp-1水平的相关性。
结论与展望
硫化氢主要通过胱硫醚-γ-裂解酶(CSE)在心肌中产生,具有心脏保护作用。前期的研究表明,雌激素可以增强雌性大鼠心肌中CSE的表达。本研究旨在探讨雌激素调节CSE表达的机制,特别是阐明雌激素受体亚型的作用以及负责雌激素作用的转录因子。研究发现CSE抑制剂或CSE小干扰RNA减弱了17β-雌二醇(E2)对原代培养的新生儿心肌细胞中H2O2-和缺氧/复氧引起的损伤的保护作用。E2在体外培养的心肌细胞和体内雌性小鼠的心肌中通过雌激素受体(ER)-α刺激CSE表达。在大鼠CSE启动子中鉴定出特异性蛋白1(Sp-1)共有位点,并发现该位点介导E2诱导的CSE表达。E2增加卵巢切除大鼠心肌中ERα和Sp-1的表达并抑制microRNA(miR)-22的表达。在原代心肌细胞中,E2通过ERα介导的miR-22下调来刺激Sp-1表达。已证实ERα和Sp-1都是miR-22的靶标。在卵巢切除大鼠的心肌中,miR-22的水平与CSE、ERα、Sp-1和抗氧化生物标志物呈负相关,与氧化生物标志物呈正相关。使用Unisense微呼吸测量系统,研究人员能够准确地测量和分析硫化氢的产生,这对于理解雌激素对心脏的保护机制以及CSE在其中的作用至关重要。总之本研究表明,雌激素通过ERα介导的心肌细胞miR-22下调来刺激Sp-1,导致CSE上调,进而导致抗氧化防御增强。ERα、miR-22和Sp-1的相互作用可能在雌性大鼠心肌氧化应激状态的控制中发挥关键作用。
